
Key Concepts
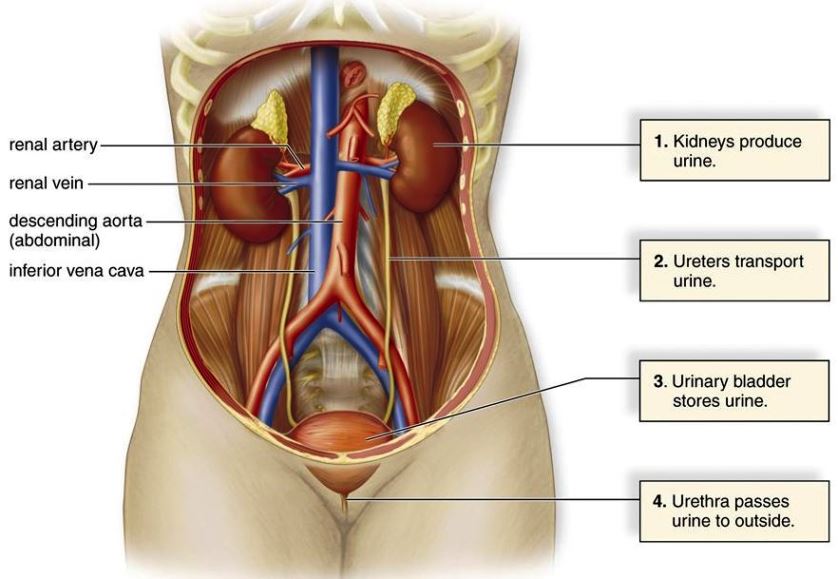
The system consisting of the kidneys, urinary ducts, and bladder. The urinary system (Fig. 1) functions in the elaboration and excretion of urine in vertebrates. Similarities are not particularly evident among the many and varied types of excretory organs found in vertebrates. The variations that are encountered are undoubtedly related to problems with which vertebrates have experienced in adapting to different environmental conditions. See also: Adaptation (biology); Excretion; Kidney; Urinary bladder; Urinary tract disorders; Urine; Urogenital system; Vertebrata
Comparative anatomy
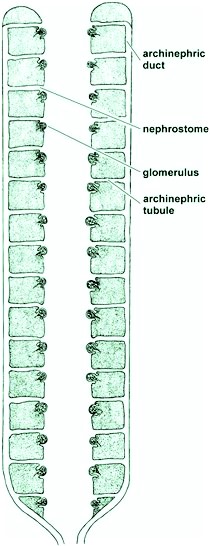
It is generally believed that the primitive vertebrate ancestor possessed an excretory organ referred to as an archinephros or holonephros (Fig. 2). This probably consisted of a pair of dorsally located ducts extending the length of the body cavity. Each duct was joined by a series of segmentally arranged tubules, with one pair to each segment. The other end of each tubule opened into the body cavity by a ciliated, funnel-shaped aperture. Close to each opening was a small knot of arterial blood vessels called an external glomerulus. From this type of kidney with its archinephric duct, the various kidneys of forms living today may originally have been derived. The larval form of the hagfish and the larvae of the caecilians (wormlike, legless amphibians of the order Apoda) are present-day vertebrates possessing kidneys of this type. See also: Apoda; Jawless vertebrates
Anamniote kidneys
The anterior portion of the archinephric kidney persists only in the adult stage of the hagfish and of certain teleost fishes, in which it is called the head kidney or pronephros. It appears in the embryos of most vertebrates as a transitory structure that usually degenerates soon after it has formed. The remainder of the kidney posterior to the pronephros is known as the opisthonephros. The opisthonephros serves as the adult kidney in lampreys, most fishes, and amphibians.
Fishes
There is much variation in shape of the opisthonephric kidneys of fishes, but they are fundamentally similar in structure. In some, they extend the length of the coelom; in others, they are short and may show various degrees of fusion. Some marine teleosts lack glomeruli and thus possess aglomerular kidneys. In elasmobranchs, the anterior ends of the kidneys of the male have been appropriated by the reproductive system. In teleost fishes, there is no connection between the testes and the opisthonephric kidneys. The posterior ends of the archinephric ducts of female fishes enter a common urinary sinus inside a small urinary papilla. The latter enters the cloaca in elasmobranchs and dipnoans; in contrast, in most other fishes, it opens directly to the outside because a cloaca is absent. See also: Elasmobranchii; Reproductive system; Teleostei
Amphibians
Adult amphibians possess an opisthonephros. The kidneys of the tailed amphibians are much like those of elasmobranch fishes, with the anterior ends in males being concerned with genital rather than urinary functions. In frogs and toads, the opisthonephric kidneys lie toward the posterior part of the abdominal cavity; the kidneys of females are not related to the reproductive system, but an intimate connection exists in males. A thin-walled urinary bladder opens into the amphibian cloaca. It has no connection with the archinephric ducts. See also: Amphibia
Amniote kidneys
In reptiles, birds, and mammals, three types of kidneys are usually recognized—the pronephros, mesonephros, and metanephros. These appear in succession during embryonic development, but only the metanephros persists in the adult. The mesonephros and metanephros actually represent different levels of the opisthonephros of lower forms, with the metanephros being equivalent to the posterior portion.
The anteriorly located pronephros appears during very early development, but it soon degenerates and the more posterior mesonephros then develops. However, the duct of the pronephros persists to become the duct of the mesonephros. This is actually the same as the archinephric duct, but is usually referred to as the Wolffian duct.
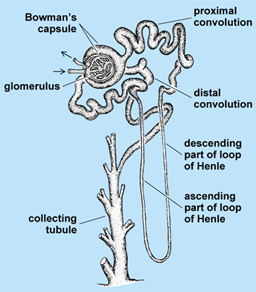
The mesonephros persists for a time and then degenerates. In the meantime, the metanephros (Fig. 3) has begun to develop from the region posterior to the mesonephros. A few mesonephric tubules and the Wolffian duct persist to contribute to the reproductive system of the male or to remain as vestigial structures.
Reptiles
The metanephric kidneys of reptiles lie in the posterior part of the abdominal cavity, usually in the pelvic region. They are small, compact, and often markedly lobulated. The posterior portion on each side is somewhat narrower. In some lizards, the hind parts may even fuse. The degree of symmetry varies, being most divergent in snakes and limbless lizards, which have notably long, narrow, lobulated kidneys in correlation with the shape of the body. One kidney may be entirely behind the other. See also: Reptilia
Snakes and crocodilians lack a urinary bladder; however, most lizards and turtles have well-developed bladders, which open into the cloaca. Except in turtles, the ureters open independently into the cloaca. In turtles, they connect to the bladder.
Birds
The kidneys of birds are situated in the pelvic region of the body cavity; their posterior ends are usually joined. They are lobulated structures with short ureters that open independently into the cloaca. Except for the ostrich, birds lack urinary bladders. Urinary wastes, chiefly in the form of semisolid uric acid, are eliminated through the cloaca along with feces. See also: Aves
Mammals
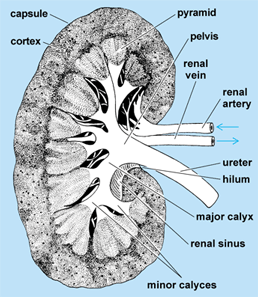
The typical mammalian metanephric kidney (Fig. 4) is a compact, bean-shaped organ attached to the dorsal body wall outside the peritoneum. The ureter leaves the medial side at a depression, termed the hilum. At this point, a renal vein also leaves the kidney, and a renal artery and nerves enter it. The kidney is surrounded by a capsule of connective tissue, under which lies the cortex. The renal corpuscles and the greater part of the secretory tubules lie entirely in the cortex. The portion of the kidney surrounded by the cortex is the medulla. It is partly composed of large areas, called the renal pyramids. The outer borders of the pyramids are divided into smaller units called lobules. The collecting tubules lie within the pyramids, but may extend well up into the cortex. The inner portion of each pyramid, in the form of a blunt papilla, projects into an outpocketing of the pelvis known as a minor calyx. Several minor calyces join together to enter major calyces, which in turn open into the renal pelvis. The renal pelvis leads to the ureter, which empties into the bladder (except in monotremes, in which it enters the urethra). Urine, which is stored temporarily in the bladder, passes to the outside through the urethra. In males, the urethra opens at the tip of the penis. In females, the condition varies, but it usually enters a vestibule, which is the terminal part of the genital tract. The kidneys of mammals are markedly lobulated in the embryo; in many forms, this condition is retained throughout life. See also: Mammalia
Urinary bladder
The kidney, or nephros, of vertebrates consists of many individual structural and functional units known as nephrons. In all known vertebrates, the nephrons empty into a common drainage (nephric) duct. At or near the posterior ends of the nephric ducts, there frequently is a reservoir for urine. This is the urinary bladder.
There are two basic varieties of bladders in vertebrates. One is found in fishes, and the reservoir is no more than an enlargement of the posterior end of each urinary duct. Frequently, the urinary ducts are conjoined, and a small bladder is formed by expansion of the common duct. The far more common type of bladder is that exhibited by tetrapods. This is a sac that originates embryonically as an outgrowth from the ventral side of the cloaca. Present in all embryonic life, it is exhibited differentially in adults. All amphibians retain the bladder, but it is lacking in snakes, crocodilians, and a few lizards; in addition, birds, with one or two exceptions, lack a bladder. It is present in all mammals. See also: Tetrapoda
Among mammals, only a few primitive forms retain a cloaca as adults. In all other mammals, it is modified in such a way as to be eliminated. Concomitantly, the openings of the excretory ducts are shifted, and the urinary bladder is established.
Physiology
Urine is produced by individual renal nephron units, which are fundamentally similar from fish to mammals (Fig. 3). However, the basic structural and functional pattern of these nephrons varies among representatives of the vertebrate classes in accordance with changing environmental demands. Kidneys serve the general function of maintaining the chemical and physical constancy of blood and other body fluids. The most striking modifications are associated particularly with the relative amounts of water made available to the animal. Alterations in degrees of glomerular development, in the structural complexity of renal tubules, and in the architectural disposition of the various nephrons in relation to one another within the kidneys may represent adaptations made to either conserve or eliminate water.
Certain basic principles underlie excretory processes in all animals. These are regulation of volume, electrolyte balance, movement of water across cell membranes, and elimination of nitrogenous substances.
Regulation of volume
Paleontological evidence strongly suggests that early vertebrates evolved in freshwater, or at least that progenitors of modern fishes had a long history of dwelling in freshwater. Kidneys of living vertebrates bear the imprint of this early evolutionary history. Except for the primitive marine cyclostome Myxine, all modern vertebrates, whether marine, freshwater, or terrestrial, have concentrations of salt in their blood that are only one-third or one-half that of seawater. The early development of the glomerulus can be viewed as a device responding to the need for regulating the volume of body fluids. Hence, in a hypotonic freshwater environment, the osmotic influx of water through gills and other permeable body surfaces would be kept in balance by a simple autoregulatory system; that is, a rising volume of blood results in increased hydrostatic pressure, which in turn elevates the rate of glomerular filtration. Similar devices are also found in freshwater invertebrates. See also: Osmoregulatory mechanisms
Electrolyte balance
A system that regulates volume by producing an ultrafiltrate of blood plasma must conserve inorganic ions and other essential plasma constituents. The salt-conserving operation appears to be a primary function of the renal tubules that encapsulate the glomerulus. As the filtrate passes along their length toward the exterior, inorganic electrolytes are extracted from them through highly specific active cellular resorptive processes that restore plasma constituents to the circulatory system.
Movement of water
Concentration gradients of water are attained across cells of renal tubules by water following the active movement of salt or other solutes. Where water is free to follow the active resorption of sodium and covering anions, as in the proximal tubule, an isosmotic condition prevails. Where water is not free to follow salt, as in the distal segment in the absence of antidiuretic hormone, a hypotonic tubular fluid results.
Nitrogenous end products
Of the major categories of organic foodstuffs, end products of carbohydrate and lipid metabolism are easily eliminated mainly in the form of carbon dioxide and water. Proteins, however, are more difficult to eliminate because the primary derivative of their metabolism, that is, ammonia, is a relatively toxic compound. For animals living in an aquatic environment, ammonia can be eliminated rapidly by simple diffusion through the gills. However, when ammonia is not free to diffuse into an effectively limitless aquatic environment, its toxicity presents a problem, particularly to embryos of terrestrial forms that develop wholly within tightly encapsulated eggshells or cases. For these forms, the detoxication of ammonia is an indispensable requirement for survival. During evolution of the vertebrates, two energy-dependent biosynthetic pathways arose that incorporated potentially toxic ammonia into urea and uric acid molecules, respectively. Both of these compounds are relatively harmless, even in high concentrations; urea, though, needs a relatively large amount of water to ensure its elimination, and uric acid requires a specific energy-demanding tubular secretory process to ensure its efficient excretion. See also: Ammonia; Nitrogen excretion; Urea; Uric acid
Urine concentration
The unique functional feature of the mammalian kidney is its ability to concentrate urine. Human urine can have four times the osmotic concentration of plasma, and some desert rats that survive on a diet of seeds without drinking any water have urine/plasma concentration ratios as high as 17. More aquatic forms, such as the beaver, have correspondingly poor concentrating ability.
The concentration operation depends on the existence of a decreasing gradient of solute concentration that extends from the tips of the papillae in the inner medulla of the kidney outward toward the cortex. The high concentration of medullary solute is achieved by a double hairpin countercurrent multiplier system, which is powered by the active removal of salt from urine while it traverses the ascending limb of Henle's loop (Fig. 5). The salt is redelivered to the tip of the medulla after it has diffused back into the descending limb of Henle's loop. In this way, a hypertonic condition is established in fluid surrounding the terminations of the collecting ducts. Urine is concentrated by an entirely passive process as water leaves the lumen of collecting ducts to come into equilibrium with the hypertonic fluid surrounding its terminations.
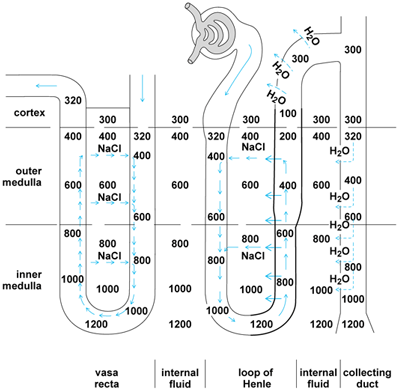
Another countercurrent exchange system operating in the straight blood vessels (vasa recta) that parallel the loops of Henle prevents washout of the hypertonic interstitial fluid of the inner medulla, where the blood is in osmotic equilibrium with the surrounding interstitial fluid. Because of the hairpin arrangement of the vasa recta, salt and other solutes are trapped and kept from being carried away from the medulla (left side of Fig. 5). As solute diffusing from the ascending capillary limb comes into equilibrium with more dilute interstitial fluid nearer the cortex, it enters the descending vessel to be redelivered to the inner medulla. This entirely passive process involving exchange of solute is the same as that operating in countercurrent heat exchangers.
Removal of most of the salt from the glomerular filtrate takes place in the proximal convoluted tubules of the renal cortex. Sodium is actively resorbed from tubular fluid, and chloride then passively follows down an electrical gradient. Removal of solute effects the simultaneous isosmotic entry of water into the cortical interstitial fluid. This solution is then rapidly restored to the general circulation by blood perfusing the cortical capillaries.

