
Key Concepts
The sense of sight, which perceives the form, color, size, movement, and distance of objects. Vision is the sensory ability allowing the qualities of an object that constitute its appearance to be perceived (Fig. 1). Of all the senses, vision provides the most detailed and extensive information about the environment. In the higher animals, especially the birds and primates, the eyes and the visual areas of the central nervous system have developed a size and complexity far beyond the other sensory systems. Visual impairments can range from relatively minor abnormalities of visual acuity (the ability to see fine details of an object) to complete blindness. See also: Binocular vision disorders; Central nervous system; Color vision; Eye (invertebrate); Eye (vertebrate); Perception; Primate color vision; Sensation; Sense organ; Vertebrate eye development; Visual impairment
Visual stimuli
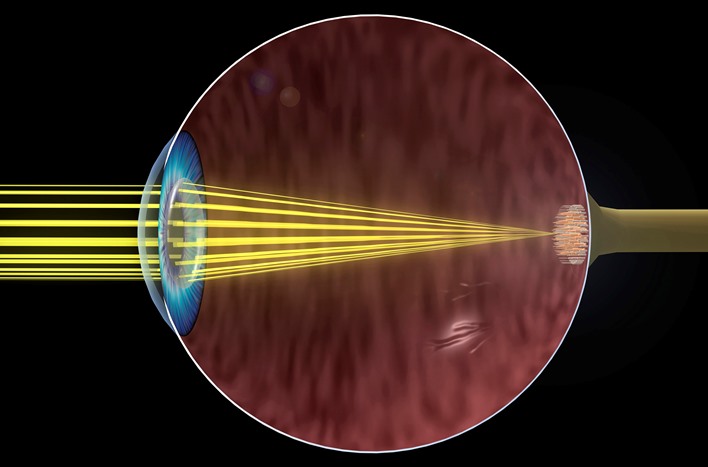
Visual stimuli are typically rays of light entering the eyes and forming images on the retina at the back of the eyeball (Fig. 2). The intensity and wavelength characteristics of the light vary according to the light source and the object from which they are reflected. Human vision is most sensitive for light comprising the visible spectrum in the range of 380–720 nanometers (nm) in wavelength. Sunlight and common sources of artificial light contain substantially all wavelengths in this range, but each source has a characteristic spectral energy distribution. In general, light stimuli can be measured by physical means with respect to their energy, dominant wavelength, and spectral purity. These three physical aspects of the light are closely related to the perceived brightness, hue, and saturation, respectively. See also: Color; Light; Optics
Anatomical basis for vision
The anatomical structures involved in vision include the eyes, optic nerves and tracts, optic thalamus, primary visual cortex, and higher visual areas of the brain. The eyes are motor organs as well as sensory; that is, each eye can turn directly toward an object to inspect it. The two eyes are coordinated in their inspection of objects, and they are able to converge for near objects and diverge for far ones. Each eye can also regulate the shape of its crystalline lens to focus the rays from the object and to form a sharp image on the retina. Furthermore, the eyes can regulate the amount of light reaching the sensitive cells on the retina by contracting and expanding the pupil of the iris. These motor responses of the eyes are examples of involuntary actions that are controlled by various reflex pathways within the brain. See also: Brain
The process of seeing begins when light passes through the eye and is absorbed by the photoreceptors of the retina. These cells are activated by the light in such a way that electrical potentials are generated. These potentials are probably responsible for many features of the electroretinogram, which is an electrical response wave that can be detected by means of electrodes attached to the outside of the eye. Impulses emerge from the eye in the form of repetitive discharges in the fibers of the optic nerve. However, the optic nerve impulses do not mirror exactly the excitation of the photoreceptors by light. Complex interactions within the retina serve to enhance certain responses and to suppress others. Furthermore, each eye contains more than a hundred times as many photoreceptors as optic nerve fibers. Thus, it would appear that much of the integrative action of the visual system has already occurred within the retina before the brain has begun to act.
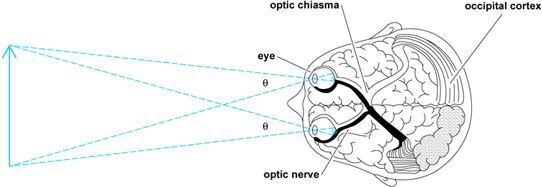
The optic nerves from the two eyes traverse the optic chiasma. Figure 2 shows that the fibers from the inner (nasal) half of each retina cross over to the opposite side; in contrast, those fibers from the outer (temporal) half do not cross over, but remain on the same side. The effect of this arrangement is that the right visual field, which stimulates the left half of each retina, activates the left half of the thalamus and visual cortex. Conversely, the left visual field affects the right half of the brain. This situation is similar to that of other sensory and motor projection systems in which the left side of the body is represented by the right side of the brain, and vice versa.
The visual cortex includes a projection area in the occipital lobe of each hemisphere of the brain. Here, there appears to be a point-for-point correspondence between the retina of each eye and the cortex. Thus, the cortex contains a "map" or projection area, wherein each point represents a point on the retina and therefore a point in visual space as seen by each eye. Other important features of an object, such as its color, motion, orientation, and shape, are simultaneously perceived. The two retinal maps are merged to form the cortical projection area. This merger allows the separate images from the two eyes to interact with each other in stereoscopic vision, binocular color mixture, and other phenomena. In addition to the projection areas on the right and left halves of the cortex, there are visual association areas and other brain regions that are involved in vision. Complex visual acts, such as form recognition, movement perception, and reading, are believed to depend on widespread cortical activity beyond that of the projection areas. See also: Stereoscopy
Scotopic and photopic vision
Night animals, such as the cat or the owl, have eyes that are specialized for seeing with a minimum of light. This type of vision is called scotopic. Day animals, such as the horned toad, ground squirrel, or pigeon, have predominantly photopic vision. Animals with photopic vision require much more light for seeing, but their daytime vision is specialized for quick and accurate perception of fine details of color, form, and texture, and location of objects. Color vision, when it is present, is also a property of the photopic system. Human vision is duplex; humans are in the fortunate position of having both photopic and scotopic vision. Some of the chief characteristics of human scotopic and photopic vision are listed in the table.
|
Characteristic |
Scotopic vision |
Photopic vision |
|---|---|---|
|
Photochemical substance |
Rhodopsin |
Cone pigments |
|
Receptor cells |
Rods |
Cones |
|
Speed of adaptation |
Slow (30 min or more) |
Rapid (8 min or less) |
|
Color discrimination |
No |
Yes |
|
Region of retina |
Periphery |
Center |
|
Spatial summation |
Much |
Little |
|
Visual acuity |
Low |
High |
|
Number of receptors per eye |
120,000,000 |
7,000,000 |
|
Cortical representation |
Small |
Large |
|
Spectral sensitivity peak |
505 nm |
555 nm |
Scotopic vision
Scotopic vision occurs when the rod receptors of the eye are stimulated by light. The outer limbs of the rods contain a photosensitive substance known as visual purple or rhodopsin. This substance is bleached away by the action of strong light so that the scotopic system is virtually blind in the daytime. In darkness, however, the rhodopsin is regenerated by restorative reactions based on the transport of vitamin A to the retina by the blood. For example, one experiences a temporary blindness upon walking indoors on a bright day, especially into a dark room or dimly lighted theater. As the eyes become accustomed to the dim light, the scotopic system gradually begins to function. This process is known as dark adaptation. Complete dark adaptation is a slow process during which the rhodopsin is restored in the rod receptors of the retina. Faulty dark adaptation or night blindness is found in persons who lack rod receptors or have a dietary deficiency in vitamin A. See also: Vitamin A
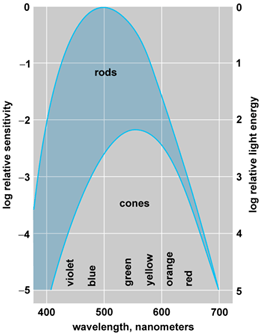
The variation of the scotopic threshold with wavelength of light is shown in the rod curve of Fig. 3. In spite of the variations in wavelength, the subject does not see any color when the intensity of the light is low enough to fall in the rod portion of the diagram. This scotopic vision is colorless or achromatic.
Normal photopic vision
Normal photopic vision has the characteristics enumerated in the table. Emphasis is placed on the fovea centralis, which is a small region at the very center of the retina of each eye.
Foveal vision
Foveal vision is achieved by looking directly at objects in the daytime. In Fig. 4, the image of a small object at F falls within a region almost exclusively populated by cone receptors. These are so closely packed together in the central fovea that their density is about 130,000 per square millimeter. Furthermore, each of the cones in the fovea is provided with a series of specialized nerve cells that process the incoming pattern of stimulation and convey it to the cortical projection area. In this way, the cortex is supplied with superbly detailed information about any pattern of light that falls within the fovea centralis.
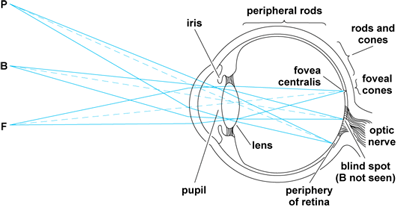
Peripheral vision
Peripheral vision takes place outside the fovea centralis (Fig. 4). As an example, look directly at a single letter at the center of a printed page. This letter and a few letters immediately adjacent to it appear clear and black because they are seen with foveal vision. The rest of the page is a blur in which the lines of print are seen as gray streaks. This is an example of peripheral vision. Vision actually extends out to more than 90° from the center, so one can detect moving objects approaching from either side. This extreme peripheral vision is comparable to night vision in that it is devoid of sharpness and color.
There is a simple anatomical explanation for the clarity of foveal vision as compared with peripheral vision. The cone receptors become less and less numerous in the retinal zones that are more and more remote from the fovea. In the extreme periphery, there are scarcely any cones, and even the rod receptors are more sparsely distributed. Furthermore, the plentiful neural connections from the foveal cones are replaced in the periphery by network connections in which hundreds of receptors may activate a single optic nerve fiber. This mass action is favorable for the detection of large or dim stimuli in the periphery or at night; however, it is unfavorable for visual acuity and color vision, which both require the brain to differentiate between signals arriving from closely adjacent cone receptors.
Space and time perception
Spatial and temporal effects are clearly apparent in the sense of sight. These two effects enable the individual to be oriented with regard to space and time in the surrounding world, and they are especially important for the perception of motion and distance.
Space perception
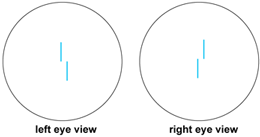
Elementary forms of space perception are vernier and stereoscopic discrimination. Here, the eye is required to judge the relative position of one object in relation to another (Fig. 5). The left eye, for example, sees the lower line in Fig. 5 as displaced slightly to the right of the upper. This is known as vernier discrimination. The eye is able to distinguish fantastically small displacements of this kind, including a few seconds of arc under favorable conditions. If the right eye is presented with similar lines that are oppositely displaced, then the images for the two eyes appear fused into one and the subject sees the lower line as nearer than the upper. This is the principle of the stereoscope. Again, it is true that displacements of a few seconds of arc are clearly seen, and they are seen as changes in distance. The distance judgment is made not at the level of the retina, but at the cortex, where the spatial patterns from the separate eyes are fused together. The fineness of vernier and stereoscopic discrimination transcends that of the retinal mosaic and suggests that some averaging mechanism must be operating in space or time (or both).
The spatial aspects of the visual field are also of interest. Good acuity is restricted to a narrowly defined region populated by densely packed cones at the center of the visual field. Farther out, in these peripheral regions, area and intensity are reciprocally related for all small sizes of stimulus field. A stimulus patch of unit area, for example, looks the same as a patch of twice the same area and half the luminance. This high degree of areal summation is achieved by the convergence of hundreds of rod receptors upon each single optic nerve fiber. It is the basis for the ability of the dark-adapted eye to detect large objects even on a dark night.
In daytime vision, spatial inhibition, rather than summation, is most noticeable. The phenomenon of simultaneous contrast is present at a border between fields of different color or luminance. This has the obvious effect of heightening contours and making forms more noticeable against their background (Fig. 6).

Time perception
The temporal characteristics of vision are revealed by studying the responses of the eye to various temporal patterns of stimulation. When a light is first turned on, there is a vigorous burst of nerve impulses that travel from the eye to the brain. Continued illumination results in fewer and fewer impulses as the eye adapts itself to the given level of illumination. Turning the light off elicits another strong neural response.
The strength of a visual stimulus depends upon its duration as well as its intensity. Below a certain critical duration, the product of duration and intensity is found to be constant for threshold stimulation. A flash of light lasting only a few milliseconds may stimulate the eye quite strongly, provided that its luminance is sufficiently high. A light of twice the original duration will be as detectable as the first if it is given half the original luminance.
Voluntary eye movements enable the eyes to roam over the surface of an object of inspection. In reading, for example, the eyes typically make four to seven fixational pauses along each line of print, with short jerky motions between pauses. An individual's vision typically takes place during the pauses, so that one's awareness of the whole object is the result of integrating these separate impressions over time.
A flickering light is one that is going on and off (or undergoing lesser changes in intensity) as a function of time. At a sufficiently high flash rate [called the critical frequency of fusion (cff)], the eye fails to detect the flicker, and the light pulses seem to fuse to form a steady light that cannot be distinguished from a continuous light that has the same total energy per unit of time. As the flash rate is reduced below the critical frequency of fusion, flicker becomes noticeable; moreover, at very low rates, the light may appear more conspicuous than flashes occurring at higher frequency. The critical frequency of fusion is often used clinically to indicate a person's visual function as influenced by drugs, fatigue, or disease. See also: Psychology

