
Key Concepts
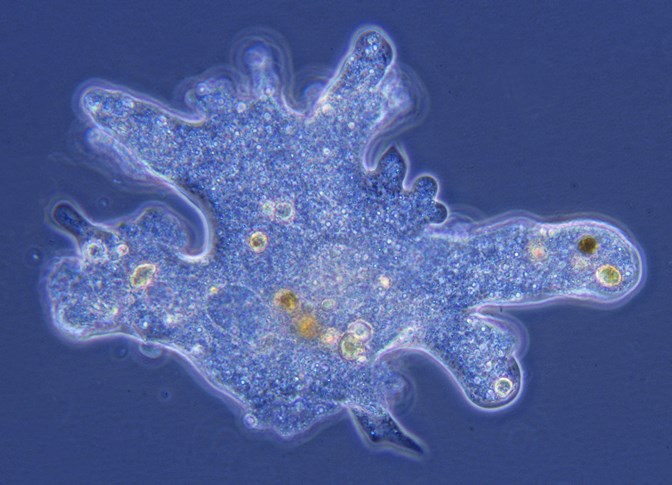
An informal grouping of single-celled eukaryotic microorganisms. In previous taxonomic schemes, certain unicellular eukaryotic microorganisms were assigned to an animal kingdom known as Protozoa. However, modern-day classifications consider this term to be obsolete and invalid, and the word protozoa (or protozoans) is used commonly as a descriptive term (Fig. 1). Moreover, although members of the informal protozoan grouping often display animal-like characteristics, they are not classified as animals, and they are not considered to be plants or fungi either. Instead, protozoa or protozoans are animal-like protists (eukaryotes that are not animals, plants, or fungi; other protists include algae, water molds, and slime molds). See also: Animal; Biological classification; Eukaryota; Protista; Taxonomy
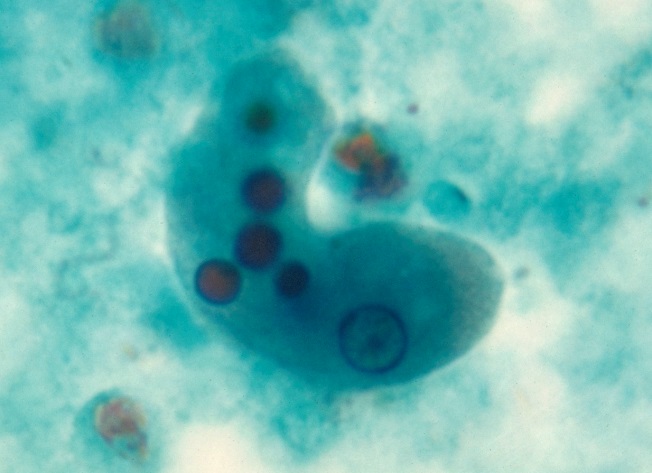
Protozoans are almost as widely distributed as bacteria, and they can be free-living or parasitic. Free-living types occur in soil, wet sand, and fresh, brackish, and salt waters. Protozoans of the soil and sand live in films of moisture on the particles. Habitats of endoparasites vary. Some are intracellular, such as malarial parasites in vertebrates. Other parasites, such as Entamoeba histolytica (Fig. 2), invade tissues, but not individual cells. Most trypanosomes live in the blood plasma of vertebrate hosts. Many other parasites live in the lumen of the digestive tract or sometimes in coelomic cavities of invertebrates, as do certain gregarines. See also: Coccidia; Gregarinia; Trypanosomatidae
Many protozoans are uninucleate (possessing a single nucleus), whereas others are binucleate or multinucleate. In addition, the number of nuclei may vary at different stages in a life cycle. Size ranges from less than 3 micrometers (µm) to a number of centimeters. Colonies are known in flagellates, ciliates, and sarcodine ameboids. Although marked differentiation of reproductive and somatic zooids (independent individuals of colonial animals) characterizes certain colonies, protozoa have not developed tissues and organs. See also: Ameba; Cell nucleus
Morphology
A protozoan may be a plastic organism (ameboid type), but changes in form are often restricted by the pellicle (a thin protective membrane). In Trypanosoma lewisi, parallel microtubules beneath the pellicle may support this layer and may also, as probably contractile structures, cause the typical undulations of these flagellates. Although protozoan form varies considerably, there is a tendency toward universal symmetry in floating species and radial symmetry in sessile types. Bilateral symmetry is rare, and most swimmers show appreciable spiral torsion.
A protective layer is often secreted outside the pellicle, although the pellicle itself may be strengthened by incorporation of minerals. Secreted coverings may fit closely—for example, cellulose-containing thecae, which are somewhat analogous to cell walls in higher plants. Thecae also may be composed of plates arranged in a specific pattern. In other protozoans, tests may be composed mostly of inorganic material, although organic (chitinous) tests occur in certain species. Siliceous skeletons, often elaborate, characterize some protozoans. A vase-shaped lorica (rigid case), from which the anterior part of the organism or its appendages may be extended, occurs in certain flagellates and ciliates. Certain marine ciliates are actively swimming loricate forms.
Colonies and aggregates
An important feature of colonies is the material that holds the component zooids together. In spheroid colonies, a matrix is secreted during development of the colony. Spheroid colonies of a few radiolarians (for example, Collozoum), unusually large examples, may measure 4–6 cm (1.6–2.4 in.) in width. In arboroid colonies, a branching pattern is produced by a branching stalk or by attachment of loricae to one another. Dispersal of such colonial species sometimes involves migratory stages. In certain spheroid colonies, reproductive and somatic zooids are differentiated. More often, members of a colony are similar. In addition to colonial types, certain protozoa may form aggregates by not separating promptly after fission. Palmella stages, analogous to spheroid colonies, are aggregates of nonflagellated organisms. See also: Radiolaria
Flagella
Flagella occur in the active or flagellated stages of various protozoans. A flagellum consists of a sheath enclosing a matrix in which an axoneme extends from the cytoplasm to the flagellar tip. In certain groups, the sheath shows lateral fibrils (mastigonemes), which increase surface area and also may modify direction of the thrust effective locomotion. The base (blepharoplast) is considered a self-replicating structure from which new flagella arise in fission. Flagellar number ranges from one or two (for most free-living species) to many (as seen on certain symbionts of termites and wood roaches). Most flagella extend forward in swimming; others extend posteriorly, sometimes driving the body forward, sometimes serving as a ski in gliding, or forming the margin of an undulating membrane. Certain organelles, as the kinetoplast, axostyle, parabasal body, or sometimes other structures, may be attached to or adjoin the blepharoplasts. In general, flagellar movements include planar sinusoidal waves, helical waves, and lateroposterior strokes. See also: Cilia and flagella
Cilia
Although typically shorter than flagella, cilia are similar in structure. Multiplicity of cilia entails complex fibrillar systems. It has been suggested that several sets of fibrils help support the cortex and ciliary system. A more delicate fibril, or tubule, running along each row of basal granules (as seen in Tetrahymena) seemingly offers little support and may function in coordination. Other ciliates, such as hypotrichs, with compound organelles, may have quite complex fibrillar systems. Compound ciliary organelles represent fused bundles or rows of cilia, with the basal granules detectable in basal plates of the organelles. Cirri, typically ventral, are fused tufts of cilia and are important in locomotion. See also: Ciliophora
Pseudopodia
Two major types have been described—the contraction-hydraulic type and the two-way flow type. Contraction-hydraulic pseudopodia are lobopodia with rounded tips and ectoplasm denser than endoplasm. The larger ones commonly contain granular endoplasm and clear ectoplasm. According to the contraction-hydraulic theory, pseudopodia arise as a result of pressure from the contracting ectoplasm on the endoplasm, forcing the latter forward, away from the posterior contraction. Local weakening of the ectoplasm results in a bulge, or developing pseudopodium. Endoplasm reaching the tip of the growing pseudopodium is diverted to the ectoplasm and converted into gel, thus becoming part of the ectoplasm. In the meantime, ectoplasm at the other end is being converted into endoplasm, which flows forward again under pressure from the ectoplasm.
Two-way flow pseudopodia include reticulopodia, filoreticulopodia, and axopodia. In all these types, cytoplasmic flow in opposite directions occurs on opposite sides of a pseudopodium.
Trichocysts
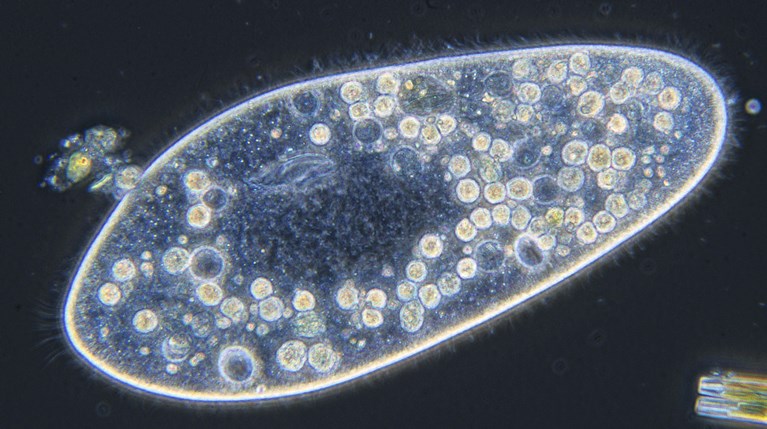
At maturity, trichocysts (minute structures that release filamentous or fibrillar threads when discharged) lie in the cortex of certain ciliates and flagellates. These structures probably serve in secretion of cyst walls and similar materials. Fibrillar trichocysts of Paramecium (Fig. 3) may be extruded partially to anchor a feeding ciliate. In addition, toxicysts of various protozoans may induce paralysis of prey. See also: Paramecium
Inclusions
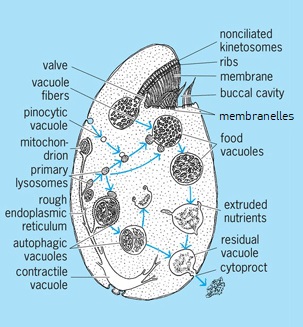
Nuclei, food vacuoles (gastrioles), chromatophores (which contain chlorophylls), stigmas (which possibly function in orientation with relation to a light source), and water-elimination vesicles are found in the cytoplasm of protozoans. In addition, the cytoplasm may contain mitochondria, Golgi material, pinocytotic vacuoles, stored food materials, an endoplasmic reticulum, and sometimes pigments of various kinds. Small vacuoles (0.5–1.0 µm) in Tetrahymena pyriformis (Fig. 4) are considered lysosomes containing acid hydrolases. See also: Cell organization; Chromatophore; Endoplasmic reticulum; Golgi apparatus; Lysosome; Mitochondria; Vacuole
Nutrition
In protozoan feeding, either phagotrophic (holozoic) or saprozoic (osmotrophic) methods predominate in particular species. In addition, chlorophyll-bearing flagellates profit from photosynthesis; in fact, certain species have not been grown in darkness and may be obligate phototrophs. See also: Nutrition; Photosynthesis
Phagotrophic feeding
Ingestion of food, followed by digestion in vacuoles, is characteristic of ameboids, ciliates, and many flagellates. Digestion follows synthesis of appropriate enzymes and their transportation to the food vacuole. Ingestion of nonnutritive latex particles also leads to acid-phosphatase activity of the food vacuoles. Primary lysosomes, which can fuse with the food vacuole, may transport hydrolases from the rough endoplasmic reticulum, as suggested for Tetrahymena (Fig. 4). Some parasitic alveolates, including sporozoans (organisms whose adult stage is not motile), are known to ingest host proteins or sometimes tissue cells or portions of them. Details of ingestion vary. Formation of food cups, or gulletlike invaginations, to enclose prey is common in ameboid organisms, many flagellates, and at least a few sporozoans. Entrapment in a sticky reticulopodial net occurs in ameboids. Phagotrophic ciliates and a few flagellates have a persistent cytostome and gullet. Many ciliates (Fig. 4) have buccal organelles (membranes, membranelles, and closely set rows of cilia) arranged to drive particles to the cytostome. Particles pass through the cytostome into the cytopharynx (gullet), at the base of which food vacuoles (gastrioles) are formed. Digestion occurs in such vacuoles. An additional feature, reported in Paramecium and Tetrahymena, is entanglement of particles in mucus secreted by the peristomial cortex or buccal cavity. The effect is a clumping of particles into larger aggregates. See also: Enzyme
Saprozoic feeding
By definition, saprozoic feeding involves passage of dissolved foods through the cortex. It is uncertain to what extent diffusion is responsible, but enzymatic activities presumably are involved in uptake of various simple sugars, acetate, and butyrate, as well as potassium and phosphate ions. In addition, external factors (for example, the pH of the medium) may strongly influence uptake of fatty acids and phosphate.
Pinocytosis
Pinocytosis resembles phagocytosis in miniature, but with ingestion apparently limited to materials in solution. First recognized in Amoeba proteus, the process has since been described in other amebas, certain ciliates, and some sporozoans. Pinocytosis is stimulated by "inducers," such as proteins, certain salts, and toluidine blue (a basic dye). In the process, small vacuoles are formed at the base of a tubular invagination from a surface, either cortex or the cytopharynx wall. The vacuoles drift into the endoplasm and mingle with other inclusions. In certain ciliates, a similar process yields secondary vacuoles from a food vacuole. In certain malarial parasites, but not in others, such pinocytosis precedes completion of digestion. See also: Endocytosis; Phagocytosis
Nutritional requirements
In nature, nutritional requirements are met by the available diet, which includes other microorganisms and materials dissolved in the external medium. Certain mineral materials, including calcium, silicon, and strontium salts, are needed for tests and other skeletal structures, and metal ions are needed in traces for enzyme systems. Nitrogen requirements of many phytoflagellates are met by an ammonium salt or by either a nitrate or an ammonium salt. The only carbon source needed by certain chlorophyll-bearing flagellates is carbon dioxide (CO2). Organic substances, such as vitamins, are needed in traces. Some few phytoflagellates are known to synthesize all required vitamins. Other protozoa need one or more vitamins and sometimes other growth factors. See also: Carbon dioxide; Vitamin
Reproduction
Reproduction occurs after a period of growth that ranges, in different species, from less than half a day to several months. General reproductive methods include binary fission, budding, plasmotomy, and schizogony. Fission, involving nuclear division and replication of organelles, yields two organisms similar in size. Budding (either internal or external) produces two organisms—one smaller than the other. In plasmotomy, a multinucleate organism divides into several daughter cells, with each containing a number of nuclei. Schizogony, which is characteristic of sporozoans, follows repeated nuclear division, yielding many uninucleate buds. In addition, reproduction includes formation of new organelles, either by division of old structures or by neoformation that is sometimes preceded by resorption of an old organelle. Blepharoplasts and basal granules, in particular, seem to be self-replicating. Formation of a new mouth (stomatogenesis) also occurs in ciliates.
Life cycles
Simple life cycles include a cyst and an active (trophic) stage undergoing growth and reproduction. In certain free-living and parasitic species, no cyst is developed. Dimorphic cycles show two active stages [for example, adult and larva; flagellate and ameba; flagellate and palmella (nonflagellated); and ameba and plasmodium], whereas polymorphic cycles show several active stages.
Encystment
Encystment involves resorption of locomotor and feeding organelles, loss of water, often rounding up of the body, and secretion of one or more cyst membranes. The wall usually is composed of secreted materials (for example, proteins, carbohydrates, and keratin), although foreign particles may be cemented into a wall. Precystic activities may include storage of reserves. Protective cysts, with fairly thick walls, often resist desiccation. In fact, dried cysts of certain amebas and flagellates have remained viable for almost 50 years in dry soil samples. Within reproductive cysts, different reproductive methods occur, including fission, budding or schizogony, gametogenesis, and syngamy. The usually thin-walled reproductive cysts afford relatively little protection.
Excystment involves absorption of water leading to rupture of cyst membranes and, when necessary, development of organelles for feeding and locomotion.
Sexual activities
Sexual activities involve production of haploid (gametic) nuclei and their fusion into a synkaryon (zygotic nucleus). Meiosis occurs at different stages in life cycles. In haploid organisms (for example, certain flagellates, coccidian parasites, malarial parasites, and gregarines), the zygote is the only diploid stage. Meiosis occurs as the zygotic nucleus divides. Diploid cycles, in which meiosis occurs during gametogenesis, are found in ciliates. See also: Meiosis
Syngamy, autogamy, and conjugation are common sexual activities. Syngamy involves fusion of two gametes, which may be morphologically similar (isogamy) or different (anisogamy). Autogamy, which is observed in certain ciliates and a few parasitic flagellates, occurs within a single organism; haploid nuclei are formed and subsequently fuse into a synkaryon. Conjugation involves pairing of ciliates, apparently following appearance of specific protein-containing substances at the surface, including cilia.
Parasitic protozoans
Parasites occur in all major groups of protozoans. Sporozoans are exclusively parasitic, as are some flagellates (for example, trichomonads and hypermastigids) and several ciliates (for example, apostomes). Various other protozoan groups contain both parasitic and free-living types. Protozoans also serve as hosts of other protozoans, certain bacteria, fungi, and algae. Many protozoan symbionts, called commensals, are more or less harmless. Cellulose-digesting flagellates of termites and ciliates of ruminants are symbiotic in the sense that they benefit their hosts. In addition to helpful digestive activities, rumen ciliates are potential food for the herbivores. See also: Parasitology
Relatively few parasites are distinctly pathogenic, causing amebiasis, visceral leishmaniasis (kala azar), African sleeping sickness (African trypanosomiasis), Chagas disease (American trypanosomiasis), malaria, tick fever of cattle, dourine of horses, and other diseases. Harmful effects may be produced in different ways. Tissue cells are invaded and destroyed in Chagas disease, leishmaniasis, coccidiosis, and malaria. Tissues may be destroyed, as in abscesses and ulcers involving Entamoeba histolytica. Transfer may involve direct migration of parasites living in aquatic hosts; direct contact (osculatory or venereal); ingestion with contaminated food or drink; and vector transfer, as in malaria or Chagas disease. See also: Malaria; Medical parasitology; Trypanosomiasis

