
The genealogical history of organisms, both living and extinct. Phylogeny represents the historical pattern of relationships among organisms which has resulted from the actions of many different evolutionary processes. Phylogenetic relationships are depicted by branching diagrams called cladograms, or phylogenetic trees. Cladograms show relative affinities of groups of organisms called taxa. Such groups of organisms have some genealogical unity, and are given a taxonomic rank such as species, genera, families, or orders. For example, two species of cats—say, the lion (Panthera leo) and the tiger (Panthera tigris)—are more closely related to each other than either is to the gray wolf (Canis latrans). The family including all cats, Felidae, is more closely related to the family including all dogs, Canidae, than either is to the family that includes giraffes, Giraffidae. The lion and tiger, and the Felidae and Canidae, are called sister taxa because of their close relationship relative to the gray wolf, or to the Giraffidae, respectively.
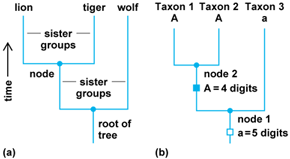
Cladograms thus depict a hierarchy of relationships among a group of taxa (Fig. 1a). Branch points, or nodes, of a cladogram represent hypothetical common ancestors (not specific real ancestors), and the branches connect descendant sister taxa. If the taxa being considered are species, nodes are taken to signify speciation events. The goal of the science of cladistics, or phylogenetic analysis, is to discover these sister-group (cladistic) relationships and to identify what are termed monophyletic groups—two or more taxa postulated to have a single, common origin.
Methodology
The acceptance of a cladogram depends on the empirical evidence that supports it relative to alternative hypotheses of relationship for those same taxa. Evidence for or against alternative phylogenetic hypotheses comes from the comparative study of the characteristics of those taxa. Similarities and differences are determined by comparison of the anatomical, behavioral, physiological, or molecular [such as deoxyribonucleic acid (DNA) sequences] attributes among the taxa. A statement that two features in two or more taxa are similar and thus constitute a shared character is, in essence, a preliminary hypothesis that they are homologous; that is, the taxa inherited the specific form of the feature from their common ancestor. However, not all similarities are homologs; some are developed independently through convergent or parallel evolution, and although they may be similar in appearance, they had different histories and thus are not really the same feature. In cladistic theory, shared homologous similarities are either primitive (plesiomorphic condition) or derived (apomorphic condition), whereas nonhomologous similarities are termed homoplasies (or sometimes, parallelisms or convergences). This distinction over concepts and terminology is important because only derived characters constitute evidence that groups are actually related.
As evolutionary lineages diversify, some characters will become modified. Examples include the enlargement of forelimbs or the loss of digits on the hand. Thus, during evolution the foot of a mammal might transform from a primitive condition of having five digits to a derived form with only four digits (Fig. 1b). Following branching at node 1, the foot in one lineage undergoes an evolutionary modification involving the loss of a digit (expressed as character state A). A subsequent branching event then produced taxa 1 and 2, which inherited that derived character. The lineage leading to taxon 3, however, retained the primitive condition of five digits (character state a). The presence of the shared derived character, A, is called a synapomorphy, and identifies taxa 1 and 2 as being more closely related to each other than either is to taxon 3. Distinguishing between the primitive and derived conditions of a character within a group of taxa (the ingroup) is usually accomplished by comparisons to groups postulated to have more distant relationships (outgroups). Character states that are present in ingroups but not outgroups are postulated to be derived. To determine relationships among the species of cats (family Felidae), comparisons to characters in other carnivore outgroups such as dogs (Canidae) or bears (Ursidae) would help distinguish primitive and derived conditions within cats.
Systematists have developed computer programs that attempt to identify shared derived characters (synapomorphies) and, at the same time, use them to construct the best phylogenetic trees for the available data. Conceptually, the two procedures—determining the polarity of character transformations and finding the best phylogenetic tree—are logically related (Fig. 2). For example, three alternative phylogenetic trees are possible for three taxa. A comparative analysis of the three might reveal that they share four characters, A--D, each of which has two alternative states. Comparisons to one or more outgroup taxa help the systematist interpret the polarity of these character states (in this example, uppercase letters are taken to be the derived conditions). Given the distributions of the character states shown in Fig. 2, postulated character transformations can be assigned to the branches of each alternative tree. In all three trees, the derived condition, A, unites the three ingroup taxa but cannot help choose among the three alternatives for their interrelationships. When the second character is examined, it is seen that the postulated derived condition, B, unites taxa 2 and 3 to the exclusion of taxon 1, which has the primitive condition, a. The derived conditions of the third and fourth characters are each found in one taxon only: C in taxon 3 and D in taxon 2, respectively. Thus, only the transformation from b to B carries phylogenetic information. In addition, tree 1 also better explains the distributions of the different character states because it requires that b transform to B only once, whereas trees 2 and 3 require two independent origins of B from b. Thus, tree 1 is the simplest hypothesis given the available data and is to be preferred. Similarly, computer programs can process data sets having large numbers of taxa and characters, examine all possible character transformations on a set of trees, and then choose that tree which accounts for the fewest number of changes.
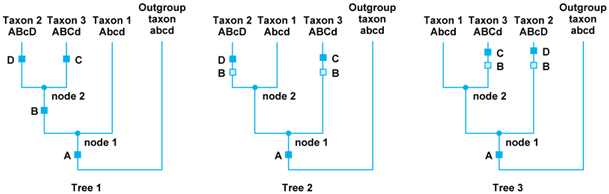
The interpretation of the status of a character shared among taxa depends upon an assessment of its history on a given tree. Thus, in tree 1 the derived character B is interpreted as a synapomorphy (it unites taxa 2 and 3); that is, B is inferred to be homologous in the two taxa. Within the framework of threes 2 and 3, however, the derived character B is interpreted as having evolved independently in taxa 2 and 3. In this case, character state B is interpreted as being homoplasious (parallel) and thus not homologous.
Value of phylogenies
Understanding how species and groups of species are interrelated is fundamental to interpreting observations across all of biology. Knowledge of phylogenetic relationships provides the basis for classifying organisms. Ideally, all taxonomic groups—genera, families, orders, and so on—should be monophyletic. Some groups, such as birds and mammals, are monophyletic; that is, phylogenetic analysis suggests they are all more closely related to each other than to other vertebrates. However, other traditional groups, such as reptiles, have been demonstrated to be nonmonophyletic (some so-called reptiles, such as dinosaurs and their relatives, are more closely related to birds than they are to other reptiles such as snakes). A major task of the science of systematics is to search for monophyletic groups. Classifications based on monophyletic groups are termed natural classifications.
Phylogenies are also essential for understanding the distributional history, or biogeography, of organisms. Knowing how organisms are related to one another helps the biogeographer to decipher relationships among areas and to reconstruct the spatial histories of groups and their biotas. See also: Biogeography
Biologists use phylogenies to make predictions about the characteristics of organisms that are still poorly studied. If one species produces a particular compound that has useful pharmacological properties, it is probable that its close relatives will share an identical or similar compound. The same is true for other structural, behavioral, or physiological characters.
History of life
Life has diversified on Earth for about 4 billion years. Systematists have described nearly 1.4 million living species, most of which are arthropods (insects, spiders, and their relatives), as well as tens of thousands of fossil species. It is estimated that from 3 million to perhaps as many as 100 million living species remain to be discovered, and the number of unknown fossil species is no doubt equally vast.
Much knowledge about the history of life has been derived from studies of comparative anatomy. Advances in understanding the history life have been made by using information derived from comparisons of DNA sequences, which largely involves diversification within the microbial world. It is often believed that fossils represent the direct ancestors of other fossils or of living organisms and, therefore, that fossils are necessary to reconstruct phylogeny. Because of the vast panoply of taxonomic and structural diversity seen throughout deep evolutionary time, reconstructing its history is a complex undertaking, and many aspects are still not well understood. Many major groups of organisms appear to have diversified in an evolutionary burst about 600 million years ago. Rapid radiations, especially those taking place long ago, pose particular difficulties for reconstructing phylogeny, for often relatively few morphological and molecular characters evolve between branching events that are so close to one another in time. However, there has been an upsurge in interest in reconstructing phylogeny, so knowledge of the history of life should improve substantially. See also: Animal evolution; Systematics; Taxonomic categories

