
Key Concepts
The processes of biological and organic change in animals by which descendants come to differ from their ancestors. Animal evolution encompasses the theory that modern animals are the modified descendants of animals that formerly existed and that these earlier forms descended from still earlier and different organisms. The concepts of evolutionary biology and their relationship to phylogeny (the genealogical history of organisms, both living and extinct), adaptation, and speciation were formulated by Charles Darwin. Much of Darwin's conclusions emanated from his investigative study of animals. Animals are multicellular organisms that feed by ingestion of other organisms or their products, being unable to derive energy through photosynthesis or chemosynthesis. Animals are currently classed into approximately 36 phyla, and each of these phyla has evolved a distinctive body plan or architecture. Representatives of the major animal groups are depicted in Fig. 1. See also: Adaptation (biology); Animal; Animal kingdom; Charles Darwin and his theory of evolution; Evolution; Macroevolution; Phylogeny; Speciation; Species concept
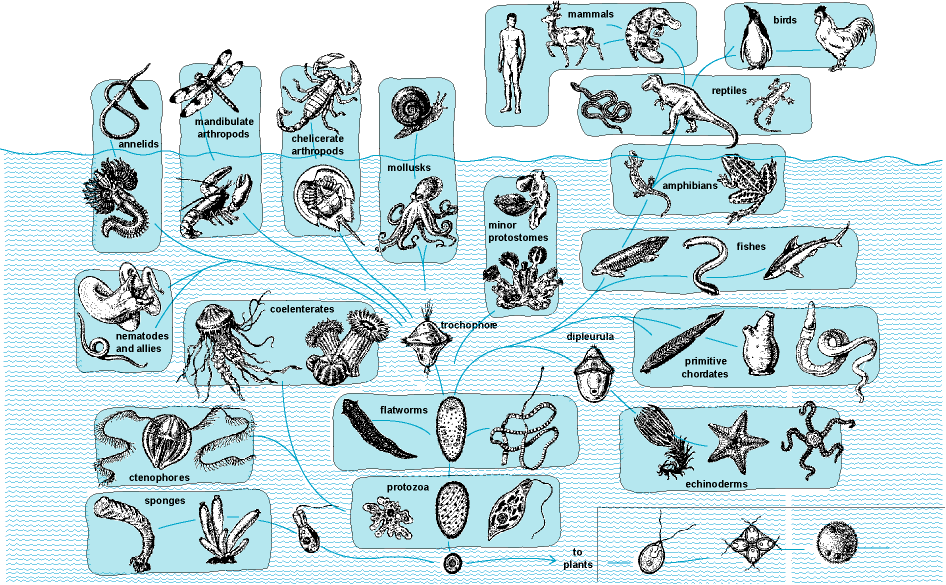
Interrelationships of animal groups
All phyla began as invertebrates, but lineages of the phylum Chordata developed the internal skeletal armature, with spinal column, which was exploited in numerous fish groups and which eventually gave rise to terrestrial vertebrates. The number of phyla is uncertain partly because most of the branching patterns and the ancestral body plans from which putative phyla have arisen are not yet known. For example, arthropods (including crustaceans and insects) may have all diversified from a common ancestor that was a primitive arthropod, in which case they may be grouped into a single phylum; or several arthropod groups may have evolved independently from nonarthropod ancestors, in which case each such group must be considered a separate phylum. So far as known, all animal phyla began in the sea. See also: Arthropoda; Chordata; Invertebrate phylogeny; Vertebrata
Several lines of evidence bear significantly upon the ancestry of these animals and of their major subdivisions. Classically, animals were grouped according to the similarities of their adult body plans; however, although the body plans have provided a basis for the recognition of most of the phyla and other distinctive animal groups, they have not provided definitive evidence of the interrelations of the phyla. A second approach is to compare patterns of early development, which should be more conservative than adult features; such comparisons have permitted the grouping of some phyla into likely alliances. Hypotheses as to the body plans of phylum ancestors have been erected from these morphologic data. It has not proven possible to corroborate any hypotheses, however, because the earliest members of the phyla (and of invertebrate classes) appear abruptly in the fossil record, and their ancestors cannot be traced. Furthermore, numbers of extinct phyla or other major animal groups also appear in the fossil record, adding branches to the tree of life that must be reckoned with, but with no definitive indication of either their origins or branching patterns. Finally, genetic changes that have accumulated after branching events separated the major groups should have left a pattern of variation in the structure of genes that may be reconstructed to yield a phylogeny. If genetic changes have occurred in a reasonably regular manner, then the amount of divergence in the structure of genes and gene products, such as protein and ribonucleic acid (RNA) molecules, should be proportional to the time since branching occurred. Some genes change so fast that their pattern of divergence is useful only for recent branching events, whereas others are so conservative that they are used to study divergences that occurred billions of years ago. These methods are promising when used in conjunction with other evidence. See also: Fossil; Gene; Taphonomy
History of major animal groups
The simplest living phyla may have no direct relationships to other animals (Fig. 2), but they indicate the versatility of multicellular design. For example, members of the Porifera (sponges) contain several differentiated cell types that form body walls, but their architecture is limited to folding these walls into chambers that are sometimes highly convoluted. Members of the Cnidaria (also termed Coelenterata, including sea anemones, corals, and jellyfish) have two cellular body layers, separated by a gelatinous layer; the inner tissue forms a digestive cavity. See also: Cnidaria; Porifera
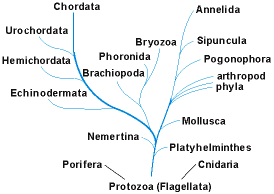
Flatworms (Platyhelminthes) and their allies have solid bodies, with both external (ectoderm) and internal (endoderm) tissue layers separated by a third cellular layer (mesoderm); there is a gut lined with endoderm. All phylogenetic evidence suggests that a flatwormlike form gave rise to the remaining phyla, which possess body cavities (other than a gut) in which reproductive and other organs can be sequestered. In about 15 of these phyla, this cavity (the coelom) is lined by mesoderm and usually functions as a hydrostatic skeleton. See also: Platyhelminthes
The coelomates constitute all of the familiar, larger-bodied animals and are commonly grouped into two main divisions (Fig. 2). In one group, the body was originally segmented, as in annelid worms or arthropods, although the segmentation may be reduced or lost in some branches. In the other group, the coelom is divided into two or three regions; these include the Enchinodermata (starfish and sea urchins), Tunicata (sea squirts), and Chordata (including humans). Additionally, there are coelomate phyla that cannot yet be assigned to either of these groups; some, such as the Brachiopoda, Phoronida, and Bryozoa, display an enigmatic mixture of traits that otherwise characterize one or the other of the main groups. In about 10 phyla (not shown in Fig. 2), the body cavity is not a true coelom, but lies between endoderm and mesoderm and is termed a pseudocoel. These pseudocoelomates are soft-bodied, small to minute, and commonly parasitic; some of them have never been found as fossils. See also: Annelida; Brachiopoda; Bryozoa; Echinodermata; Phoronida; Tunicata (Urochordata)
Some features of the cells of primitive animals resemble those of the single-celled Protozoa, especially the flagellates, which have long been believed to be animal ancestors. Molecular phylogenies have supported this idea and also suggest that the phylum Cnidaria arose separately from all other phyla that have been studied by this technique. Thus, animals may have evolved at least twice from organisms that are not themselves animals, thereby representing a grade of evolution and not a single branch (clade) of the tree of life. Sponges also have been suspected of an independent origin, and it is possible that some of the extinct fossil phyla arose independently or branched from sponges or cnidarians. See also: Protozoa
The earliest undoubted animal fossils (the Ediacaran fauna) are soft-bodied, and they first appear in marine sediments nearly 650 million years ago. This fauna lasted about 50 million years and consisted chiefly of cnidarians or cnidarian-grade forms, although it contains a few enigmatic fossils that may represent groups that gave rise to more advanced phyla. Then, nearly 570 million years ago, just before and during earliest Cambrian time, a diversification of body architecture began that produced most of the living phyla, as well as many extinct groups. The body plans of some of these groups involved mineralized skeletons that created, for the first time, an extensive fossil record because these skeletons are more easily preserved than soft tissues. The earliest mineralized skeletons are chiefly minute, including the small shelly fauna of earliest Cambrian age; moreover, although some of these fossils are primitive members of living phyla, many (perhaps 10 to 20 kinds of them) are distinctive and quite possibly represent phyla or at least major branches of phyla that soon became extinct, geologically speaking. The soft-bodied groups were also markedly diversified, although their record is so spotty that their history cannot be traced in detail. A single, exceptionally preserved soft-bodied fauna from the Burgess Shale of British Columbia, which is about 530 million years old, contains not only living soft-bodied worm phyla, but extinct groups (perhaps a dozen) that cannot be placed in living phyla and do not seem to be ancestral to them. Among these is Wiwaxia, a novel form that bears spinelike elements similar to some of the small shelly fossils of Early Cambrian age, thus providing evidence that the small shelly forms did indeed include novel types. Clearly, an early radiation of animals produced a vast array of novel body plans in an evolutionary episode that has never been surpassed. See also: Burgess Shale; Cambrian; Ediacaran biota
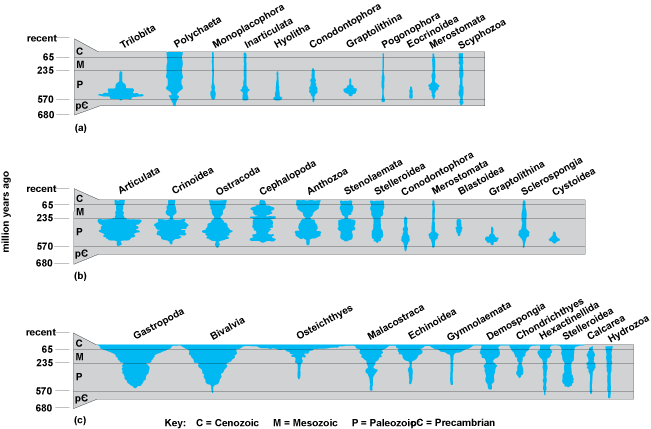
Following the early phase of rampant diversification and of some concurrent extinction of phyla and their major branches, the subsequent history of the durably skeletonized groups can be followed in a general way in the marine fossil record (Fig. 3). The composition of the fauna changed continually, but three major associations can be seen: one dominated by the arthropodlike trilobites during the early Paleozoic [panel (a) of Fig. 3], one dominated by articulate brachiopods and crinoids (Echinodermata) in the remaining Paleozoic [panel (b) of Fig. 3], and one dominated by gastropod (snail) and bivalve (clam) mollusks during the Mesozoic and Cenozoic [panel (c) of Fig. 3]. The mass extinction at the close of the Paleozoic that caused the contractions in so many groups (Fig. 3) may have extirpated more than 90% of marine species and led to a reorganization of marine community structure and composition into a modern mode. Resistance to this and other extinctions seems to have been a major factor in the rise of successive groups to dominance. On land, the fossil record is too spotty to trace in detail the invasions of the terrestrial environment, which must have been under way by the mid-Paleozoic. Annelids, arthropods, and mollusks are the more important invertebrate groups that made the transition to land. The outstanding feature of terrestrial fauna is the importance of the insects, which appeared in the late Paleozoic and later radiated to produce several million living species, surpassing all other life forms combined in this respect. See also: Cenozoic; Extinction; Extinction (paleontology); Insecta; Mesozoic; Mollusca; Paleozoic; Trilobita
Chordate origins
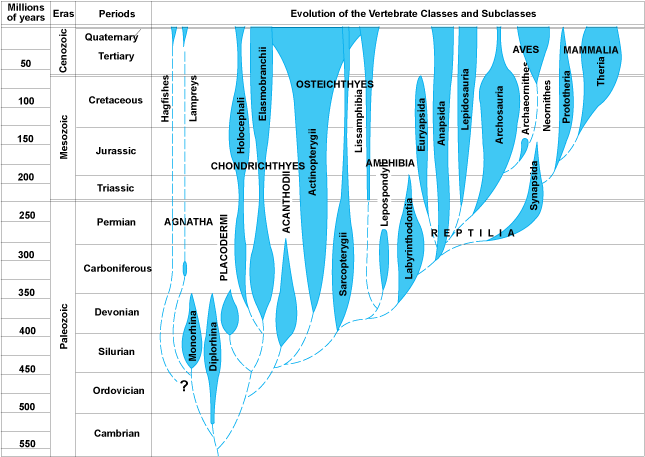
The phylum Chordata consists largely of animals with a backbone, that is, the Vertebrata, including humans (Fig. 4). The group, however, includes some primitive nonvertebrates, termed the protochordates: lancelets, tunicates, acorn worms, pterobranchs, and possibly the extinct graptolites and conodonts. The interrelationships of these forms are not well understood. With the exception of the colonial graptolites, they are soft-bodied and have only a very limited fossil record. They suggest possible links to the Echinodermata in developmental, biochemical, and morphological features. In addition, some early Paleozoic fossils, the carpoids, have been classified alternatively as chordates and as echinoderms by various investigators, again suggesting a link. In spite of these various leads, the origin of the chordates remains basically unclear. See also: Conodont; Graptolithina
Chordates are characterized by a hollow, dorsal, axial nerve chord, a ventral heart, a system of slits in the pharynx that serves variously the functions of feeding and respiration, a postanal swimming tail, and a notochord that is an elongate supporting structure lying immediately below the nerve chord. The protochordates were segmented, although sessile forms (such as the tunicates) show this only in the swimming, larval phase. Even the free-swimming forms were not truly active, and they used the gill apparatus primarily for feeding on small particles trapped on mucous streams generated in the pharynx. There are two hypotheses concerning the nature of the protochordates. One holds that all protochordates were originally sessile, although having active larvae, and that the vertebrates arose from the larval phase through increased development of the segmented muscular trunk and associated nervous system. The alternative hypothesis proposes that the sessile condition seen in many protochordates is secondary.
The first vertebrates were fishlike animals in which the pharyngeal slits formed a series of pouches that functioned as respiratory gills. An anterior specialized mouth permitted ingestion of food items large in comparison with those of the filter-feeding protochordates. Vertebrates are first known from bone fragments found in rocks of Cambrian age, but more complete remains have come from the Middle Ordovician. Innovations, related to greater musculoskeletal activity, included the origin of a supporting skeleton of cartilage and bone, a larger brain, and three pairs of cranial sense organs (nose, eyes, and ears). At first, the osseous skeleton served as protective scales in the skin, as a supplement to the notochord, and as a casing around the brain. In later vertebrates, the adult notochord is largely or wholly replaced by bone, which encloses the nerve chord to form a true backbone. All vertebrates have a heart that pumps blood through capillaries, where exchanges of gases with the external media take place. The blood contains hemoglobin in special cells that carry oxygen and carbon dioxide. During the exchanges, the oxygen content of the cells is increased and the content of carbon dioxide is decreased. In most fishes, the blood passes from the heart to the gills and thence to the brain and other parts of the body. In most tetrapods and in some fishes, blood passes to the lungs, is returned to the heart after oxygenation, and is then pumped to the various parts of the body.
Fish evolution
The jawless fish, known as Agnatha, had a sucking-rasping mouth apparatus rather than true jaws. They enjoyed great success from the Late Cambrian until the end of the Devonian. Most were heavily armored, although a few naked forms are known. They were weak swimmers and lived mostly on the bottom. The modern parasitic lampreys and deep-sea scavenging hagfish are the only surviving descendants of these early fish radiations. See also: Devonian; Jawless vertebrates
Members of a new type of vertebrate, the Gnathostomata, characterized by true jaws and teeth, arose during the Middle to Late Silurian. They constitute the great majority of fishes and all tetrapod vertebrates. The jaws are modified elements of the front parts of the gill apparatus, and the teeth are modified bony scales from the skin of the mouth. With the development of jaws, a whole new set of ecological opportunities was open to the vertebrates. Along with this, new swimming patterns appeared, made possible by the origin of paired fins, forerunners of which occur in some agnathans. See also: Gnathostomata; Silurian
Four groups of fishes quickly diversified (Fig. 2). Of these, the Placodermi and Acanthodii are extinct. Members of the Placodermi were heavily armored fishes, the dominant marine carnivores of the Silurian and Devonian, rivaled only by the large eurypterid arthropods. Members of the Acanthodii were filter-feeders, mostly of small size. They are possibly related to the dominant groups of modern fishes, that is, the largely cartilaginous Chondrichthyes (including sharks, rays, and chimaeras) and the Osteichthyes (the higher bony fishes). These also arose in the Late Silurian, but diversified later. See also: Acanthodii; Chondrichthyes; Osteichthyes; Placodermi
Conquest of land
The first land vertebrates, the Amphibia, appeared in the Late Devonian and were derived from an early group of osteichthyans called lobe-finned fishes, of which two kinds survive today: (1) the Dipnoi or lungfishes, and (2) the crossopterygian coelacanth Latimeria. They were lung-breathing fishes that lived in shallow marine waters and in swamps and marshes. The first amphibians fed and reproduced in or near the water. True land vertebrates, the Reptilia, with a modified (amniote) egg that could survive on land, probably arose in the Mississippian. See also: Amniota; Amphibia; Coelacanthiformes; Dipnoi; Mississippian
Reptile radiations
By the Middle Pennsylvanian, a massive radiation of reptiles was in process. From this radiation, one can trace the line leading to mammals as well as the lineages of the great reptiles that dominated the Mesozoic Era. The most prominent reptiles belong in the Diapsida: dinosaurs, lizards and snakes, and pterosaurs (flying reptiles). The birds, Aves, which diverged from the dinosaur radiation in the Late Triassic or Early Jurassic, are considered to be feathered dinosaurs, and thus members of the Diapsida, whereas older authorities prefer to treat them as a separate class. In addition, there were several Mesozoic radiations of marine reptiles, including ichthyosaurs and plesiosaurs. Turtles (Chelonia) first appeared in the Triassic. See also: Aves; Chelonia; Dinosauria; Jurassic; Pennsylvanian; Reptilia
Mammalian origins
The line leading to mammals can be traced to primitive Pennsylvanian reptiles, that is, the Synapsida, which diversified and spread worldwide during the Permian and Triassic. The first true mammals, based on characteristics of jaw, tooth, and ear structure, arose in the Late Triassic. Derived mammals, marsupials (Metatheria) and placentals (Eutheria), are known from the Late Cretaceous, but mammalian radiations began only in the early Cenozoic. By the end of the Eocene, all the major lines of modern mammals had become established. Mammals are easily separated into distinct groups (orders), but their relationships are not easy to discover from fossil records because of the explosion of mammalian evolution in the early Cenozoic. Molecular analyses (blood proteins, deoxyribonucleic acid, and ribonucleic acid) of living mammals show that the most primitive group of placentals is the edentates (sloths, armadillos, and anteaters). An early large radiation included the rodents, primates (including monkeys, apes, and humans), and bats, possibly all closely related to the insectivores and carnivores. The newest radiations of mammals include the elephants and sea cows, whereas the whales are related to the artiodactyls (cattle and camels). Because of biogeographic isolation, marsupials came to flourish in Australia and South America, whereas the placentals diversified widely in Eurasia and North America. See also: Cenozoic; Cretaceous; Eocene; Mammalia; Permian; Synapsida; Triassic
Primates and humans
Among the early and primitive lines of placental mammals were the Paleocene and Eocene plesiadapoids, from which the primates arose. They were mostly nocturnal, insect- and fruit-eating animals with forwardly directed eyes and a locomotor system well developed for an arboreal life. The living primates consist of prosimians; tarsiers; lemurs; monkeys and anthropoids; apes; and humans and near relatives, the hominids. During the Oligocene, monkeys diverged into Old World and New World lines. From the former, the apes arose in the late Oligocene, including the gibbons, orangutans, chimpanzees, and gorillas. See also: Apes; Oligocene; Primate
The human subspecies Homo sapiens sapiens diverged from the apes about 5 million years ago. The closest living relatives among the apes are chimpanzees and gorillas. The earliest known hominids consist of fossils of the genus Australopithecus, excavated from rocks formed about 4.2 million years ago in eastern Africa. Several distinct lines of Australopithecus, under various generic names, coexisted for a long period of time, and these australopiths also existed along with H. habilis, which lived from about 2 million to 1.75 million years ago. This early species of Homo was notable for its relatively large brain, and this species probably included the first tool user. It was superseded by H. erectus, a larger animal with a greatly increased relative brain size. About 1 million years ago, H. erectus spread from Africa to eastern and southern Asia. See also: Australopith; Fossil humans
Skeletal remains anatomically similar to those of H. sapiens have been found in northern Africa in fossil beds formed about 300,000 years ago. Truly modern forms appeared some 100,000 years ago. They penetrated Europe some 50,000 to 60,000 years ago and spread throughout the Earth, occupying the Old World first and then working over to Australia; eventually, about 15,000 to 20,000 years ago, they entered North America and spread rapidly to South America. See also: Paleontology

