
Key Concepts
The study of life history as recorded by fossil remains. Paleontology investigates the life of the past through analyses of preserved fossils and their traces (Fig. 1). The term fossil, from the Latin “fossilis” (digging; dug up), originally referred to a variety of objects dug from the Earth, some of which were believed to be supernatural substances imbued with mystical powers. However, in a modern context, fossils can be defined as recognizable remains or traces of activity of prehistoric life. This broad definition takes in a diversity of ancient remains, but specifically excludes inorganic, mineralized structures, even those that spuriously resemble life forms (for example, dendritic patterns of manganese crystals: dendrites), sometimes termed pseudofossils (false fossils). See also: Fossil; Index fossil; Trace fossils

The definition of fossils makes several qualifying statements. For example, fossils must be recognizably tied to once-living organisms, excluding amorphous organic matter such as coal and petroleum, although these are certainly derived from the products of organisms and are sometimes referred to as “fossil fuels.” The definition also encompasses two broad categories of fossils: (1) remains that are primarily skeletal hard parts (body fossils) and (2) traces of activity that are evidence of behavior of living organisms (trace fossils). The latter can be broadly construed to include tracks, trails, burrows, borings, bite and gnaw marks, fecal remains (coprolites), and gizzard stones (gastroliths) polished in the digestive activity of dinosaurs. In addition, certain organisms, especially bacteria, archaeans, algae, and vascular plants, produce distinctive and unique organic molecules, whose degraded remnants can be extracted from sediments and isolated; these biomarkers form a type of chemical fossil. See also: Paleobiochemistry
The term prehistoric is also a key part of the definition of fossils. Remains of recent animals and plants are not considered as fossils simply because they are not old enough. Paleontologists commonly use, as an arbitrary definition of “prehistoric,” the approximate end of the last glacial advance, about 10,000 years before the present (BP). In some cases, ancient organism remains that are somewhat less than 10,000 years old are termed “subfossils.” Notably, artifacts made by people are not termed fossils; instead, these artifacts constitute the data of the related science of archeology, the study of human civilizations. See also: Archeology
The fossil record thus includes a very diverse class of objects ranging from molds of microscopic bacteria in rocks more than 3 × 109 years old to unaltered bones of fossil humans in ice-age gravel beds formed only a few thousand years ago. The quality of preservation ranges from the occasional occurrence of soft parts (skin and feathers, for example) to barely decipherable impressions made by shells in soft mud that later hardened to rock. The most common fossils are hard parts of various animal groups. Therefore, the fossil record is not an accurate account of the complete spectrum of ancient life, but is biased in overrepresenting those forms with shells or skeletons. Fossilized worms are extremely rare, but it is not valid to make the supposition that worms were any less common in the geologic past than they are now.
Applications of paleontology
Paleontology lies on the boundary between two disciplines—biology and geology. The field of paleontology has been subdivided in various ways. For example, a distinction can be made between paleozoology (invertebrate and vertebrate paleontology) and paleobotany (the study of fossil plants and algae). Micropaleontology deals with those fossils that must be studied with microscopes, including compound light and scanning electron microscopes. This field unites a highly varied series of fossil groups, including shelled protists such as radiolaria and foraminifera, tiny skeletons of planktonic diatoms, and coccoliths (the latter two sometimes referred to as nannofossils in recognition of their extremely small size), as well as conodonts (minute toothlike fossils that formed parts of the feeding apparatuses in early eel-like chordates). Because of their tiny size and great abundance in small samples, microfossils have traditionally been the most economically valuable fossils; in fact, many micropaleontologists have been employed by petroleum companies to aid in the identification of particular stratigraphic levels, including those bearing petroleum. Most invertebrate paleontologists are employed in geology departments, where they are involved in a wide array of stratigraphic, paleobiological, and paleoenvironmental studies. Conversely, many vertebrate paleontologists are allied with biology or medical departments because of their studies of vertebrate skeletal anatomy and evolution. Paleobotanists are traditionally plant anatomists, employed in botany departments. Palynology involves the study of organic-walled microfossils, such as pollen and spores, but also including enigmatic marine microfossils such as acritarchs (algal resting cysts of uncertain affinities) and chitinozoans (possible egg capsules of Paleozoic organisms). However, much modern paleontology defies simple categorization and is more commonly classified along lines reflecting application of fossil data in various disciplines; these include taxonomy/systematics, evolutionary studies, biostratigraphy, taphonomy, ichnology (the study of trace fossils), and paleoecology/paleoenvironmental studies (Fig. 2). See also: Biology; Geology; Micropaleontology; Paleobotany; Palynology
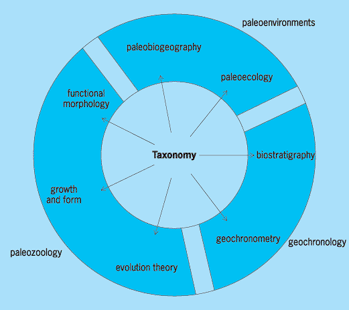
Systematics and taxonomy
The classification of fossil species into a hierarchical grouping of taxa is usually based on similarities in morphological characters. Many thousands of fossil species have been described and hundreds more are discovered and described each year; it has been estimated that the known fossil record comprises only 1–5% of the total number of easily preservable organisms that once lived on Earth. Beyond the very necessary, if mundane, description, illustration, and naming of new species lies the more challenging exercise of arranging these species into a classification that reflects evolutionary relationships or phylogeny. Several different approaches to classification have been used by paleontologists. Traditional classifications and most current ones use a modified Linnean classification, a hierarchy of nested categories beginning with the species, defined qualitatively based on overall similarities of body (or skeletal) morphology. Species in turn are grouped into related groups, genera; genera into families; families into orders; orders into classes; classes into phyla; and phyla into kingdoms. In addition, cladistics or phylogenetic systematics attempts to group taxa into dichotomous trees based upon the shared possession of unique features. Such classifications take into account a number of characters to arrive at the most parsimonious or simplest trees of relationships between taxa and their sister taxa. See also: Numerical taxonomy; Phylogeny; Speciation; Systematics; Taxonomy
Evolutionary paleontology
Increasingly, systematic paleontologists are interfacing their studies of phylogeny based on the morphology of ancient organisms with supplemental techniques, based on the biochemistry of living organisms. For example, biologists have long examined the early development of organisms or embryology for clues as to deep relationships among groups of animals. Biologists have developed techniques to determine relationships among various living taxa based on similarities of portions of their DNA or complex molecules (for example, proteins and hemoglobin). Measures of molecular distance between any two living species can give a sense of how closely related the two are, independently of their external form or morphology. Using these techniques, together with cladistics, paleobiologists and molecular biologists have come to some radical interpretations of the tree of life. Moreover, molecular biologists have found evidence that the base pairs in some strands of DNA undergo mutations in a stochastic manner, somewhat akin to radioactive decay. On this basis, they postulate that changes in DNA structure occur in a regular clocklike, time-governed manner. If so, the total amount of difference between the DNA of any two related species or other taxa can be used to calculate the amount of time that has elapsed since the two taxa diverged from a common ancestor. Thus, this molecular-clock technique provides a method, independent of paleontology, for determining the timing of evolutionary branching.
Biostratigraphy
A major task of any historical science, such as geology, is to arrange events in a time sequence and to describe them as fully as possible. Arguably the most applied field of paleontology and its traditional linkage with geology is in the field of biostratigraphy. This field employs a principle established in the early 1800s by British engineer William Smith—the principle of “faunal and floral succession,” that is, suites of particular, unique fossil species are typical of successive time intervals. Geology, the study of Earth history, did not become a modern science until the nineteenth century, when a worldwide timescale based on fossils was established; Earth history could not be deciphered until events that occurred in different places were related to one another by their position in a standard time sequence (Fig. 3). See also: Geologic time scale
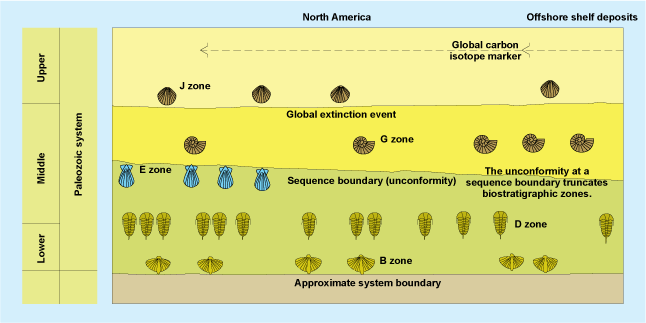
Because of evolution, there is no recurrence of species in later rocks; hence, fossils provide an arrow of time and are unambiguous markers of increments of relative time known as biozones. Certain fossils are more useful in this pursuit than others, and these form biostratigraphic guide or index fossils; these include groups that are abundant, geographically widespread, environmentally tolerant, and rapidly evolving. Although few types of organisms share all of these traits, certain groups, such as free-swimming and planktonic organisms (for example, some foraminifera, graptolites, ammonoids, and conodonts), meet most criteria and are among the most widely used zonal fossils. The primary procedures of biostratigraphy involve establishing the ranges of species and using these index fossils to correlate rocks of similar age in different locations. In some cases, the precision of correlation can be improved by using overlapping ranges of several species, or assemblages of fossils. Correlations also can be improved by methods called graphic correlation in which the first and last appearances of many species in two or more sections are plotted against one another. See also: Stratigraphy
Taphonomy
The field of taphonomy deals with the processes of fossil preservation both prior to burial (biostratinomy) or after burial (fossil diagenesis). Paleontologists pursuing studies of taphonomy may study the rates and processes of decay, disarticulation, breakage, abrasion, and disintegration of organisms and the compaction and mineralization of their remains. The objectives of taphonomy are twofold. First, by studying processes that affect potential fossil assemblages in modern times, it is possible to recognize the extent of biases of the fossil record in different environments. Second, fossils, as sedimentary particles, provide very important clues to many physical and chemical processes in ancient environments. For example, shallow-water, high-energy environments yield heavily fragmented and abraded skeletal remains, whereas muddy bottoms in deeper-water areas may preserve delicate shells and articulated crabs or sea urchins. See also: Taphonomy
Ichnology
The realm of ichnology deals with trace fossils and their interpretation. This field (together with taphonomy) interfaces closely with sedimentology because most trace fossils are in one sense literally a part of the sediments or organic sedimentary structures. Trace fossils have proven to be very useful as indicators of depositional environments. The size, depth, and diversity of traces may indicate the level of oxygen in marine bottom waters; the degree of deformation or sharpness of burrows is an indicator of sediment firmness and the degree of compaction; and distinctive trace fossil assemblages may provide a proxy for water depth. In addition, of course, traces provide a direct record of the behavior of ancient organisms and may be useful in identifying patterns of locomotion, sediment feeding, and even predator-prey relationships. See also: Sedimentology
Paleoecology and paleoenvironmental analyses
Paleoecology is the ecology of ancient organisms, that is, their modes of life and interactions with physical and biotic environments. Modes of life of ancient organisms are reconstructed on the basis of (1) biological information from closely related living analogs, (2) functional morphology (function of skeletal parts inferred from detailed studies of their form) of the fossils themselves, and (3) association with sediments or other fossils, which give environmental information (Fig. 4).
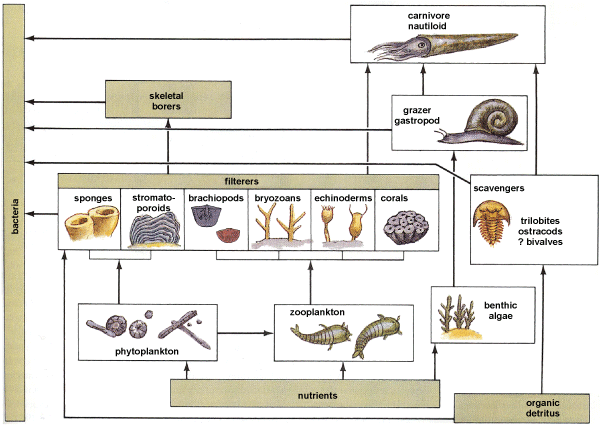
Environments are reconstructed using multiple sources of data from sedimentary rocks, including the sediments themselves (grain size, texture, and color) and sedimentary structures (for example, mud cracks and ripple marks). In some instances, particularly with relatively recent fossils, knowledge of the environmental tolerances of modern species can be used to infer the environments of the same or closely related fossil species. Other, less direct applications of fossil data may be extremely useful in inferring ancient environmental parameters. For example, the forms of coral skeletons are commonly reflective of the energy of the environment in which they grew. See also: Paleoecology
Paleoclimatology
Fossils provide very significant clues to ancient climates. The presence of growth rings in ancient woods, for example, provides evidence of seasonality, and the shapes of tree leaves prove to be a sensitive gauge of temperature—leaves with smooth margins are typical of warmer climates, whereas those with ragged or serrated margins typify cooler temperatures. The ratio of the two leaf types can be used to hindcast ancient mean annual temperatures to within 1°C. In a similar way, the coiling directions (left-hand versus right-hand) in certain planktonic foraminifera (shelled protozoans) are temperature-dependent (more left-handed coiling in cooler temperatures) and the left–right ratio can be used to hindcast temperatures accurately, at least for the last few million years.
Furthermore, the geochemistry of fossil skeletons may encode information about paleotemperatures. For example, the oxygen isotopic ratios (18O/16O) of calcium carbonate (calcite or aragonite) of normal marine shells are dependent upon the temperature at which they were secreted. Thus, the 18O/16O ratio may be utilized indirectly to reconstruct the temperature of the water from which they were secreted. See also: Paleoclimatology
Paleogeography
Fossils are indispensable guides for determining the positions of continents and seas in former times. Organisms isolated by barriers (such as deep ocean basins) evolve, although in some cases with parallel evolution to those in other areas, leading to geographically distinct provinces. The history of changing fossil assemblages provides evidence for the development and breakdown of barriers. The formation of the Isthmus of Panama can be dated, for example, by studying the distributions of marine and terrestrial fossils. Before North America and South America were connected by this land bridge, Atlantic and Pacific marine faunas were very similar, but the mammals of the two continents were completely different. However, in South American rocks deposited after the Isthmus was formed, there are fossils of North American mammals, which had migrated over the newly formed land. Likewise, Atlantic and Pacific marine faunas, isolated from each other by the rise of the Isthmus, began to evolve in different directions; this increasing difference can be traced in the fossils of successively younger rocks. Fossils also provide evidence that continental drift has occurred. If South America and Africa were once united, their faunas should be similar during that time. As they drifted apart, there should be stronger and stronger faunal differences. See also: Paleogeography
Biological aspects
The key biological aspects of paleontology support the evolutionary processes of life throughout geologic time. See also: Animal evolution; Macroevolution; Organic evolution; Plant evolution
Evolutionary process and life history
The most fundamental fact of paleontology is that organisms have changed throughout the history of Earth and that each geological period has its characteristic forms of life. When Charles Darwin published his theory of evolution in 1859, paleontological evidence for evolution was meager, but it began to accumulate quickly. The “feathered reptile” (Archaeopteryx), an almost perfect link between dinosaurs and birds, was discovered early in the 1860s, and the first fossil humans were found a few years later. In the light of this evidence and the purview of a more scientific culture, evolution was accepted as the only reasonable explanation for change in the history of life. To a paleontologist, life's outstanding attribute is its evolution (Fig. 5). See also: Archaeopteryx; Charles Darwin and his theory of evolution
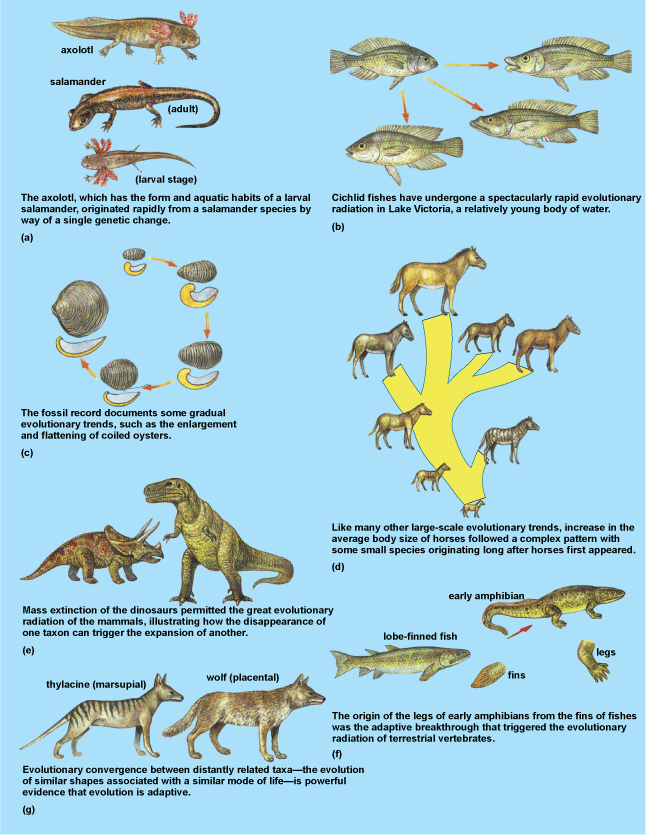
An evolutionist has two major interests—first, to know how the process of evolution works, which is accomplished largely by studying the genetics and population structure of modern organisms; second, to reconstruct the events produced by this process, that is, to trace the history of life. This is the paleontologist's exclusive domain. Any modern animal group is merely a stage, frozen at one moment in time, of a dynamic, evolving lineage. Fossils give the only direct evidence of previous stages in these lineages. Horses and rhinoceroses, for example, are very different animals today, but the fossil history of both groups is traced to a single ancestral species that lived early in the Cenozoic Era. From such evidence, a tree of life can be constructed whereby the relationships among organisms can be understood.
Life properties
Evolution is responsible for life's outstanding properties, including its diversity and its adaptation to the environment.
Diversity
A paleontologist studies diversity. Then, the number and geographic distribution of species in a more inclusive taxonomic group, such as vertebrates, can be tabulated for different periods of geologic history. When this is done for all major groups, very definite patterns emerge. Life history shows both periods of rapid diversification—evolutionary radiations—and brief intervals of rapid diversity decline—mass extinctions. These are major events in the history of life, and their causes have provided a major research agenda for paleontology. Evolutionary radiations may have resulted from new adaptive breakthroughs (for example, the evolution of legs and lungs in lobe-finned fishes), environmental triggers (for example, increased levels of atmospheric oxygen), or the availability of adaptive "niche space" (such as occurred in the Cambrian or following mass extinctions). In contrast, at the close of the Permian and Cretaceous periods, for example, extensive extinction of species occurred in many major groups at about the same time (Fig. 6). See also: Biodiversity; Extinction; Extinction (paleontology)
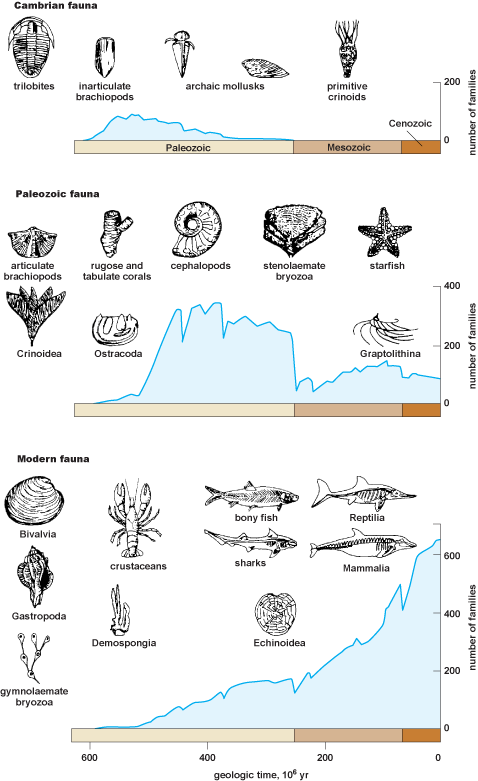
Long-term rates of evolution
The fossil record provides information on the often linked rates of evolution and extinction of various large groups of organisms. Vertebrate paleontologist George Gaylord Simpson determined the average rates of taxonomic turnover of many different taxonomic groups of animals and found that various taxa have characteristic rates of change and that certain groups evolve much more rapidly than others. Rapidly evolving taxa include mammals and ammonites; conversely, other groups such as horseshoe crabs and lingulid brachiopods evolve slowly and provide excellent examples of living fossils, that is, extant organisms that have changed very little in millions of years. Many slowly evolving groups have very generalized modes of life or live in protected environments, but just why different groups show such varying rates of change is one of the great mysteries of paleontology. See also: Living fossils
Large-scale patterns and processes
Fossils also provide very important insights into large-scale evolutionary patterns that cannot be studied with modern organisms. New species may arise by one of two processes: (1) anagenesis, or gradual change within a lineage or evolving line of populations, such as the increase in size in successively younger populations of fossil oysters (Fig. 5c); or (2) cladogenesis (speciation), or splitting of a species into two or more distinct species, typically following geographic isolation (Fig. 5d). One of the most important observations about the fossil record is that, counter to the predictions of Darwin, many species arise geologically rapidly, probably during cladogenesis, but then show stasis (little or no change) for much of their history. This staccato pattern of evolution is referred to as punctuated equilibrium. This pattern has proven to be very common, especially in many marine invertebrate animals, and it suggests that most evolutionary change occurs during short bursts associated with cladogenesis. In addition, long-term trends commonly observed in the fossil record include pattern convergence—the independent evolution of similar adaptations in distantly related groups of organisms.
Adaptation and exaptation
The term adaptation refers to the way that organic form is fitted to function (Fig. 5). According to Darwinian evolution, this fit is shaped by natural selection, actually the elimination of suboptimal features and the accumulation, by default, of mutations or changes that promote survival or reproduction. Given the raw materials of life, an engineer could scarcely design a better flying machine than a bird, yet the bird evolved naturally from reptilian ancestors, which did not fly. As Darwin realized, however, it is the imperfections and oddities of nature that provide the best insights into evolution. Some features seen in fossil and living organisms are vestigial, such as the side splint bones in horses' hoofs, which as tracked through the fossil record are evidently remnants of once functional side toes. The sources of adaptive features are also varied, but constrained by evolutionary history. Many seemingly new features may appear abruptly simply by accelerating or slowing down development of existing features, a process referred to as heterochrony. For example, simple parasitic organisms may evolve by greatly accelerating their rates of reproductive maturation; alternatively, the proportions of the adult human cranium resemble those of juvenile great apes, suggesting that it may have evolved by retention of juvenilelike features into adulthood. Both of these scenarios represent examples of paedomorphosis—adults of descendants are juvenilelike adults compared to their ancestors. See also: Adaptation (biology); Heterochrony
Other adaptations arise from preexisting structures that were present as nonfunctional side products of evolution or were initially adapted to other functions. For example, the bamboo stripping “thumbs” of pandas are actually a modified element of their wrist bones. Such exaptations are undoubtedly very common in evolution (Fig. 5f).
Paleontologists study the development of new adaptations by a variety of techniques. For example, consider the increase in brain size between apelike ancestors and modern humans. To study this, a paleontologist employs the following: (1) studying the rocks in which fossil bones are found to look for signs of any environmental change that might have made increased brain size advantageous; (2) determining the advantages of a larger brain, if possible; for example, it may have conferred, upon ancient humans, the ability to learn to control fire for warmth and cooking (charred bones of edible animals in caves inhabited by ancient humans could be used as evidence for such an assertion); and (3) applying mathematical techniques to obtain a precise measurement of the rate of evolution.
Sketch of life history
The range of life forms represented in the fossil record is extraordinarily broad, but can be subdivided into two realms—the marine and terrestrial ecosystems.
Marine ecosystem
Life on Earth probably originated in the early ocean, and possibly in mid-oceanic rifts, nearly 4 billion years ago (Fig. 7). The actual record of simple prokaryotic microbes ranges back more than 3.5 billion years. The record of these simple bacteria in the form of stromatolites (masses of layered sediments trapped on mucilaginous bacterial mats) is extensive throughout the Precambrian and later. Eukaryotes with nucleated cells and organelles evolved perhaps 2.5 billion years ago, in association with elevated levels of atmospheric oxygen; their organelles may represent symbiotic bacteria. Unicellular and multicellular algae appeared by approximately 2 billion years ago (Fig. 7). Single-celled animal-like protists (formerly, protozoans) are questionably represented by fossils about 800 million years old. Animal-like trace fossils, including simple burrows, may extend back at least 600 million years, whereas strings of fecal pellets preserved in ancient sediments indicate the presence of animals with guts. See also: Eukaryotae; Marine ecology; Marine sediments; Prokaryotae; Stromatolite
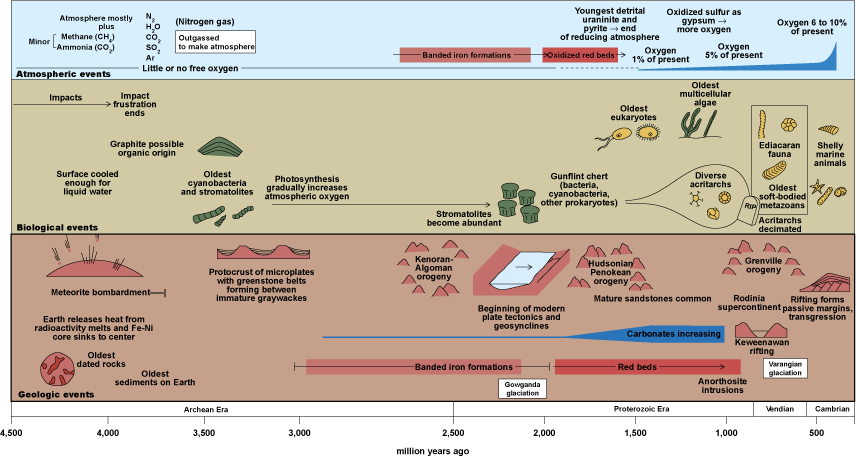
Moreover, extraordinary assemblages of large “leaflike” and radially symmetrical organisms occur as imprints in fine-grained marine sandstones worldwide about 580–550 million years ago—the Ediacaran Period of the Neoproterozoic Era. The exact affinities of these Ediacaran organisms (“vendobionts” or “vendozoans”) have been the subject of debate, with some asserting that they represent early animals such as jellyfish, sea pens (colonial marine cnidarians that resemble a quill pen), and perhaps annelid worms; others have argued that the Ediacaran organisms represent a different type of life form, possibly simple, baglike mouthless organisms that died out without leaving descendants. Regardless of the affinities of the Ediacaran organisms, the extraordinary discovery in the late 1990s of fossil animal embryos in China provides ample evidence for simple animals, such as sponges and relatives of jellyfish, nearly 600 million years ago. See also: Ediacaran biota; Vendobionts (Precambrian evolution)
The most dramatic recorded event in the history of animal life occurred about 543–530 million years ago (Early Cambrian Period)—the so-called Cambrian Explosion (Fig. 8). Within less than 10 million years, nearly all of the major body plans of life, and perhaps others that are now extinct, appeared in the oceans. These include brachiopods, mollusks, arthropods (trilobites, crustaceans, and many others), echinoderms (simple early relatives of starfish), and the chordates. A further phase of diversification in the following Ordovician Period gave rise to a much larger number of class- and order-level groupings. For example, some 20 different classes of echinoderms existed at this time, including the ancestors of crinoids, starfish, and sea urchins that have persisted to the present day.
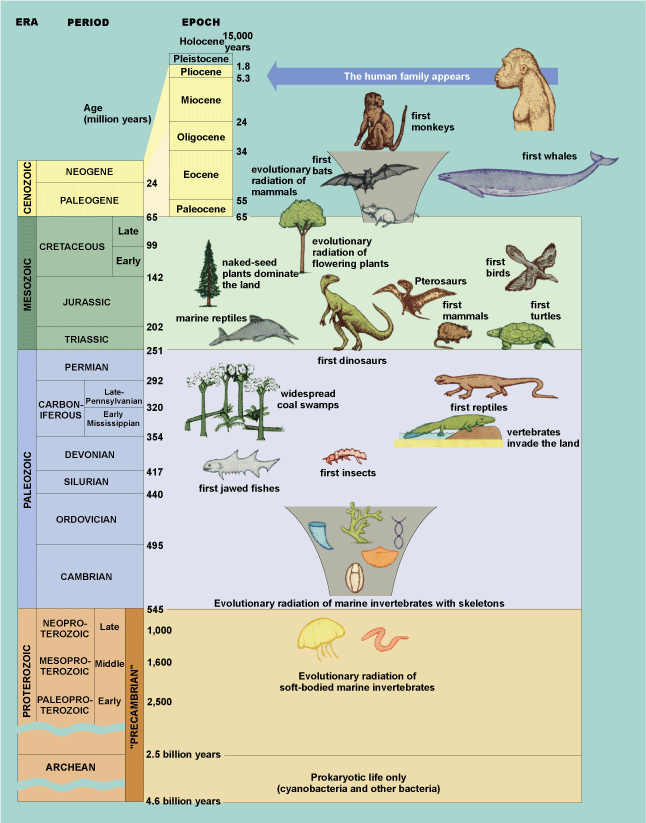
Vertebrates (animals with backbones) are now known from fossils to range back nearly as far as most invertebrates. Recent discoveries from China indicate that early vertebrates, simple jawless fishes, had evolved at least 530 million years ago. Conodont animals represent an early, eel-like vertebrate group that flourished for more than 300 million years before going extinct in the Triassic Period. Armored jawless fishes (ostracoderms) and the first jawed fishes appeared early in the Ordovician Period, with jaws evolving from gill bars. The Devonian Period (417–365 million years ago) is often termed the “Age of Fishes” because of the rapid diversification of a large number of jawed fish groups—placoderms (which diversified and then went extinct within the period), sharks, bony ray-finned fishes, and lobe-finned fishes, including lungfish and crossopterygians. See also: Origin of vertebrates
Predator-prey interactions in the marine ecosystem also intensified in two pulses. The middle Paleozoic showed rapid diversification of jawed fish predators and animals with the ability to crush and bore into shells. Potential prey organisms show a variety of probable responses, including increased shell thickness and spinosity. See also: Predator-prey interactions
During the Jurassic and Cretaceous, the marine ecosystem appears to have become increasingly “high energy,” possibly owing to an increased nutrient runoff fostered by angiosperm plant weathering of soils. The planktonic ecosystem experienced dramatic evolution, and diatoms and coccoliths became exceptionally common and formed large amounts of sediment. Radiolarians flourished in offshore settings and planktonic foraminifera appeared for the first time in the Cretaceous. This rise of primary and secondary producers was matched by unprecedented evolution of the predatory consumer guilds. A second and perhaps more intense phase of predator-prey interaction termed the “Mesozoic marine revolution” yielded highly complex predator-prey interactions involving a host of new predators. Teleost fishes, some of them huge, shared the key advance of a swim bladder for buoyancy regulation and increased maneuverability. A host of more efficient predatory sharks appeared, and these were matched by an unprecedented number of large predatory marine reptiles. Ichthyosaurs, plesiosaurs, and sea-going crocodilians flourished in the middle-to-late Mesozoic Era, and mosasaurs (giant swimming relatives of the monitor lizards) diversified in the Cretaceous. With the exception of sharks and bony fishes, most of the large predators died out near or at the end of the Cretaceous. The niches left vacant from these large reptiles were later partially filled by predaceous whales beginning in the Eocene. However, the slightly later occurring baleen whales, the most gigantic animals on Earth, feed directly on plankton, which they sieve from the water.
Terrestrial ecosystem
The terrestrial ecosystem evolved late relative to the marine ecosystem. Spore-bearing land plants such as mosses and lichens may have invaded rocky coastlines as far back as the Proterozoic; however, vascular land plants—tiny psilophytes—did not appear until the Late Ordovician or Silurian periods about 440 million years ago. These plants possessed the key adaptive breakthrough of tracheids to allow conduction of fluids up the stem, but they lacked both roots and leaves. The first land invertebrate animals, probably relatives of the millipedes, appeared at about the same time, as evidenced by trace fossils in paleosols (ancient soil deposits) from the Ordovician of Pennsylvania. Many new plant groups, including lycopods, sphenopsids (“horsetails”), ferns, and the ancestors of gymnosperms, evolved during a very critical time in land plant evolution in the Devonian Period (about 416–359 million years ago). The oldest known fossil forests, supporting trees up to 15 m (50 ft) in height, are known from stumps preserved in place in paleosols in eastern New York. Late in the Devonian (approximately 365 million years ago), one group of trees also evolved the first seeds, a reproductive breakthrough that permitted invasion of drier, upland areas. See also: Terrestrial ecosystem
Associated with the radiating land plants in the Devonian were some of the first spiders and insects. Also, at about this time, the first semiterrestrial vertebrates, the early tetrapods, ancestors of modern amphibians, evolved apparently from lobe-finned fishes or crossopterygians. These animals possessed several key adaptive breakthroughs, including fully developed lungs and tetrapod limbs with digits, effective for walking on land. See also: Tetrapoda
During the succeeding Carboniferous Period, coal swamps flourished in coastal wetlands fringing the rising Appalachian Mountains (in the eastern United States) and the corresponding Hercynian Mountains (in ancestral Europe). Coals, typically overlying heavily leached soils or underclays, were formed largely of primitive plants such as lycopods and seed-ferns. Casts of stumps of these plants at Joggins, Nova Scotia, have yielded skeletons of some of the oldest animals assignable to the reptiles. These animals possessed another key evolutionary breakthrough, amniotic eggs, with protective membranes and a shell to prevent desiccation of embryos; this feature allowed reptiles to become the first fully terrestrial animals. Drying of coal swamps during the following Permian Period led to a shift toward drier, upland vegetation, including conifers. Also, new groups of reptiles appeared, such as the mammal-like reptiles, a precursor of which included the pelycosaurs. Some, such as Dimetrodon, possessed specialized vertebral “sails,” which possibly aided in thermal regulation. The so-called mammal-like reptiles or therapsids also flourished in the Late Paleozoic. Both groups underwent extinction at the end of the Paleozoic Era. See also: Coal paleobotany; Pelycosauria; Reptilia; Therapsida
Also during the Carboniferous Period, primitive insects that were flightless evolved both wings and wing-folding ability, enabling a more fully aerial mode of life. See also: Insecta
During the Late Triassic Period, about 220 million years ago, the dinosaurs appeared. These animals dominated Mesozoic terrestrial faunas, but they coexisted with small, furry, and warm-blooded mammals, which also evolved in the Late Triassic from therapsid ancestors. Notably, flight evolved twice among the archosaurs. First, the pterosaurs (batlike reptiles that are a sister group to the dinosaurs) appeared in the Triassic; by the Cretaceous, these creatures evolved into the largest flying animals ever, with wingspans exceeding 15 m (50 ft). Second, remains of the first birds, including imprints of feathers, are found in fine-grained Solnhofen Limestone of Jurassic age from Bavaria, Germany. These animals apparently evolved from small, feathered theropod dinosaurs, which were relatives of the large predatory tyrannosaurids. See also: Archosauria; Dinosauria; Mammalia; Pterosauria
In terms of plant life, the Triassic was an age of ferns and early conifers, but the succeeding Jurassic Period is sometimes called the "Age of Cycads," a group of related gymnosperms with short, stocky trunks and palmate leaves. Later, midway through the Cretaceous Period, a very important new group of seed-bearing plants—the angiosperms or flowering plants—appeared (Fig. 8); their key breakthrough, double fertilization, led to accelerated reproduction. These plants coevolved with new groups of pollinating insects, including butterflies and moths. Dinosaurs continued to flourish during the Cretaceous and included the largest terrestrial carnivores of all time, such as Tyrannosaurus, which may have weighed 6 tons (5500 kg). In response to these predators, herbivorous dinosaurs evolved varied defenses, including armor in ankylosaurs and horns in the ceratopsians, such as Triceratops, the youngest known dinosaur. Dinosaurs, pterosaurs, and many marine animals (for example, swimming reptiles and ammonoids) became extinct at the end of the Cretaceous Period. A great debacle in the form of an asteroid impact occurred at this time, as evidenced by enrichment of iridium and shocked quartz in Cretaceous-Tertiary boundary clays about 65 million years ago, and this contributed to the demise of these organisms.
The demise of dinosaurs in terrestrial ecosystems may have opened ecological space and cleared the way for an extremely rapid adaptive radiation of mammals during the Cenozoic Era, often called the “Age of Mammals.” However, this era could equally be called the “Age of Angiosperms,” the “Age of Frogs,” the “Age of Snakes,” or the “Age of Songbirds,” as all of these groups also evolved and radiated during the Cenozoic. The evolution of C-4 plants, with their more efficient photosynthetic pathways, including the constantly growing grasses, was closely tied to the rise of ungulates, that is, hoofed, grazing mammals, during the mid-Cenozoic. These included cloven-hoofed, even-toed ungulates (pigs, camels, and cattle), as well as rhinos and horses, a group of odd-toed ungulates, whose evolution of long legs, reduced side toes, and abrasion-resistant high-crowned teeth is well documented.
Finally, about 6 million years ago, the hominid family evolved in Africa. Fossils document the early members of the genus Australopithecus, which gave rise to the larger brained genus Homo about 2 million years ago. Subsequently, Homo sapiens evolved about 200,000–100,000 years ago also in Africa, and then spread across the Old World and eventually across the Bering land bridge into North America. This immigration may have ultimately led to a wave of extinction, especially of large mammals such as mammoths and mastodons, perhaps as a result of overhunting by the one species (humans) whose activities continue to alter the terrestrial ecosystem in many ways. See also: Fossil humans

