Key Concepts
The modification of living organisms during their descent, generation by generation, from common ancestors. Evolution, also known as biological or organic evolution, encompasses the processes of change in organisms by which descendants come to differ from their ancestors, and it includes the history of the sequence of such changes (Fig. 1). Evolution includes two major processes: anagenesis, the alteration of the genetic properties of a single lineage over time; and cladogenesis, or branching, whereby a single lineage splits into two or more distinct lineages that continue to change anagenetically. Most importantly, there is no controversy about the reality of evolution as a historical event. That organisms have descended from common ancestors is accepted by knowledgeable biologists as fact. Molecular and other similarities point to the fact that all living things are related to each other by common ancestry. See also: Animal evolution; Macroevolution; Plant evolution

Study of evolution
The subject matter of evolutionary biology may be roughly divided into the analysis of the history of evolutionary events and the analysis of the mechanisms, or processes, of evolutionary change. The study of evolutionary history attempts to determine the ancestry of and genealogical relationships among different kinds of organisms; the pathways by which their morphological, biochemical, and other features have become modified; the history by which they arrived at their present geographical distributions; and the changes in the diversity and number of species throughout geological time. The methods by which such inferences are made include analysis of the fossil record (although this may be an inadequate record) and the phylogenetic analysis of living taxa. Phylogenetic analysis, using data on the comparative anatomy (Fig. 2), molecular characteristics [for example, protein and deoxyribonucleic acid (DNA) sequences], and geographical distributions of organisms, is part of the province of biological systematics. See also: Biodiversity; Biogeography; Fossil; Phylogeny; Speciation; Systematics; Taphonomy

The analysis of the mechanisms of evolutionary change addresses primarily the factors that cause changes in the genetic composition of populations and species, and those that influence diversification and extinction of species. The mathematical theory of population genetics is important to this enterprise. Experimental and observational testing of the theory includes molecular, genetic, and developmental analysis of genetic variation and the mechanisms by which it arises; ecological genetics, that is, the study of the impact of ecological factors on genetic change of populations; studies in functional morphology, physiology, behavior, and ecology that address the adaptive value of genetically different traits; and taxonomic and phylogenetic analyses that shed light on various evolutionary processes (for example, cladogenesis). Thus, the study of evolution embraces all of biology. See also: Ecology; Genetics; Population ecology; Population genetics
Historical importance of Charles Darwin
Charles Darwin (1809–1882) came to think of the possibility of evolution toward the end of his 5-year (1831–1836) voyage as naturalist on the HMS Beagle. He conceived of the theory of natural selection in 1838, and spent the next 20 years synthesizing and amassing evidence and refining his ideas, which were published in 1859 as On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. This book accomplished two things. Darwin marshals evidence from every quarter of biology and geology that evolution has in fact occurred: that living things are descended with modification from common ancestors. Second, he presents an explicit, purely mechanistic theory of the causes of evolution. Every species, Darwin points out, has hereditary variation in numerous characteristics. Some variants will be better suited to the exigencies of life than others, and hence will survive better or reproduce more prolifically than the inferior variants. Because descendants inherit their superior properties, the proportion of individuals in the population bearing superior characteristics will increase, and the proportion with inferior traits will decrease from generation to generation, until the species has been transformed. The new character itself is subject to further variation and to further alteration by this process of natural selection; thus, in the vastness of time, the feature comes to differ extremely from the original form, although it is a great change accomplished in small steps. Because different populations experience different environments and adapt to different resources, numerous forms may diverge from an original stock, with each adapted to a different environment or way of life. This branching process, continued through the immensity of geological time, gives rise to the great “tree of life.” See also: Charles Darwin and his theory of evolution
Mechanisms of species transformation
Anagenesis consists of change in the genetic basis of the features of the organisms that constitute a single species. Populations in different geographic localities are commonly considered members of the same species if they exchange members at some rate and interbreed with each other (or are thought to be potentially able to interbreed); however, unless the level of interchange (gene flow) is very high, some degree of genetic difference among different populations is likely to develop. The changes that transpire in a single population may be spread to other populations of the species by gene flow. See also: Species concept
Genetic variation
Almost every population harbors several different alleles (forms of a gene) at each of a great many of the gene loci (fixed positions of a gene); hence, many characteristics of a species are genetically variable. All genetic variations ultimately arise by mutation of the genetic material. Broadly defined, mutations include changes in the number or structure of the chromosomes and changes in individual genes, including substitutions of individual nucleotide pairs, insertion and deletion of nucleotides, and duplication of genes. Many such mutations alter the properties of the gene products [ribonucleic acid (RNA) and proteins] or the timing or tissue localization of gene action, and consequently affect various aspects of the phenotype (that is, the morphological and physiological characteristics of an organism). Whether and how a mutation is phenotypically expressed often depends on developmental (epigenetic) events. See also: Allele; Chromosome; Gene; Mutation
Natural selection
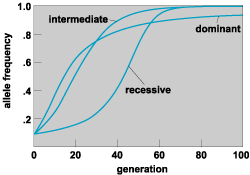
The fundamental event in evolution is a change in the frequency of an allele in a population. In its full form, this entails the spread through a population of an allele that, having just come into existence by mutation, is very rare, but which ultimately comprises 100% of the gene copies at that locus. The allele is then said to have been fixed in the population. One of the factors that causes this process is natural selection.
Natural selection is a consistent difference in the average rate at which genetically different entities in descendants are left to subsequent generations; such a difference arises from differences in fitness (that is, in the rate of survival, reproduction, or both). The entities referred to are usually different alleles at a locus, or phenotypically different classes of individuals in the population that differ in genotype. Thus, selection may occur at the level of the gene, as in the phenomenon of meiotic drive, whereby one allele predominates among the gametes produced by a heterozygote. Selection at the level of the individual organism, which is the more usual case, entails a difference in the survival and reproductive success of phenotypes that may differ (in body size, for instance) at one locus (for example, genotypes AA and aa differ in size) or at more than one locus (for example, AABBCC versus aabbcc, where different letters denote different loci and where uppercase and lowercase letters denote different alleles at a locus). As a consequence of the difference in fitness, the proportion of one or the other allele increases in subsequent generations (Fig. 3). Typically, the relative fitness of different genotypes depends on environmental conditions.
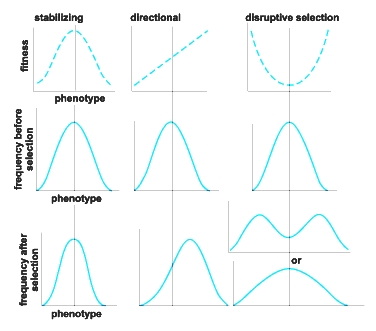
Selection is sometimes directional, meaning that one extreme state of a phenotypic feature is most fit and ultimately becomes fixed if the environmental conditions to which it is adapted prevail long enough (Fig. 4). Often, however, selection is stabilizing, meaning that an intermediate phenotype is most fit. If this phenotype is produced by a heterozygote at a single locus, selection maintains both alleles in the population. Several cases of stabilizing selection are known in which the heterozygote is most fit. Genetic variation also can be maintained by diversifying selection, whereby several different phenotypes are favored. Diversifying (disruptive) selection includes the phenomenon of frequency-dependent selection: the rarer that a genotype is, the higher does its fitness become. In addition to divergent evolution, evolution is often convergent: different organisms sometimes experience similar selection pressures and hence evolve at least superficially similar characteristics. See also: Convergent evolution
Random genetic drift
Different alleles of a gene that provides an important function do not necessarily differ in their effect on survival and reproduction; such alleles are said to be neutral. The proportion of two neutral alleles in a population fluctuates randomly from generation to generation by chance because not all individuals in the population have the same number of surviving offspring. Random fluctuations of this kind are termed random genetic drift. If different alleles do indeed differ in their effects on fitness, both genetic drift and natural selection operate simultaneously. The deterministic force of natural selection drives allele frequencies toward an equilibrium, whereas the stochastic (random) force of genetic drift brings them away from that equilibrium. The outcome for any given population depends on the relative strength of natural selection (the magnitude of differences in fitness) and of genetic drift (which depends on population size).
Speciation
The great diversity of organisms has come about because individual lineages (species) branch into separate species, which continue to diverge. This splitting process, that is, speciation, occurs when genetic differences develop between two populations that prevent them from interbreeding and forming a common gene pool. The genetically based characteristics that cause such reproductive isolation are usually termed isolating mechanisms, but there is little reason to believe that they evolve specifically to prevent interbreeding. Rather, reproductive isolation seems to develop usually as a fortuitous by-product of genetic divergence that occurs for other reasons (either by natural selection or by genetic drift).
Adaptations
A frequent consequence of natural selection is that a species comes to be dominated by individuals whose features equip them better for the environment or way of life of the species. Such features are termed adaptations. Although many features of organisms are adaptive, not all are, and it is a serious error to suppose that species are capable of attaining ideal states of adaptation. Some characteristics are likely to have developed by genetic drift rather than natural selection, and thus are not adaptations; others are side effects of adaptive features, which exist because of pleiotropy (the quality of a gene having more than one phenotypic effect) or developmental correlations. The phylogenetic history of a species determines its current state and thus the kinds of variations that may be available to natural selection. Most species are not capable of adapting ideally to all environmental changes: more than 99% of all the species that have ever lived are extinct. See also: Adaptation (biology); Extinction
Rates of evolution
The characteristics of a species evolve individually or in concert with certain other traits that are developmentally or functionally correlated. Because of this mosaic pattern of evolution, it is meaningful to speak of the rate of evolution of characters, but not of species or lineages as total entities. Therefore, in some lineages, such as the so-called living fossils, many aspects of morphology have evolved slowly since the groups first came into existence, but evolution of their DNA and amino acid sequences has proceeded at much the same rate as in other lineages. Every species, including the living fossils, is a mixture of traits that have changed little since the species' remote ancestors, and traits that have undergone some evolutionary change in the recent past. See also: Living fossils
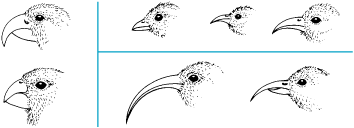
Rates of evolution are highly variable: whereas many characteristics have changed hardly at all for many millions of years in some lineages, others have responded rapidly to changes in selection pressures. Moreover, the fossil record of certain organisms, especially marine invertebrates, seems to indicate that species often change rather abruptly in morphology after being virtually static for up to a million years; the geological record, however, is usually so coarse that the “abrupt” changes may well have proceeded gradually over a period of many thousands of years. This pattern of stasis punctuated by brief periods of change has been termed punctuated equilibrium. In addition, there is abundant evidence that rates of evolution are greatest when a lineage adapts to new ecological opportunities—to vacant ecological niches. Rapid divergent evolution is common, for example, when species colonize islands that harbor few competitors; similarly, the rate of evolution is high in lineages that have survived mass extinction events. The usual pattern in such instances is one of adaptive radiation: the origin, by speciation, of numerous descendant lineages that become adapted in different ways to a variety of available resources (Fig. 5). A famous example is the radiation of Darwin's finches in the Galápagos Islands, where related species have diverged in beak morphology and have become specialized for feeding on resources that on continents are typically preoccupied by unrelated families of birds. Moreover, the major adaptive radiation seen in mammals occurred soon after the demise of the last dinosaurs, leading many investigators to suspect that the mammalian radiation was possible only because competition had been alleviated. Therefore, extinction has played an important role in the history of life, making possible the subsequent diversification of groups that otherwise might not have flourished. See also: Ecological competition; Extinction (paleontology); Island biogeography; Oceanic islands as evolutionary laboratories