
Key Concepts
The study of spatial and temporal patterns in the abundance and distribution of organisms and of the mechanisms that produce those patterns. Population ecology investigates how and why populations change over time (Fig. 1). In general, species differ dramatically in their average abundance and geographical distributions, and they display a remarkable range of dynamical patterns of abundance over time, including relative constancy, cycles, irregular fluctuations, violent outbreaks, and extinctions. Thus, the aims of population ecology are threefold: (1) to elucidate general principles explaining dynamic population patterns; (2) to integrate these principles with mechanistic models and evolutionary interpretations of individual life-history tactics, physiology, and behavior, as well as with theories of community and ecosystem dynamics; and (3) to apply these principles to the management and conservation of natural populations. See also: Conservation of species; Ecological modeling; Ecology; Mathematical ecology; Theoretical ecology
Definition of a population
A population is the total number of individuals of a given biological species found in one place at one time. In practice, ecologists often deal with density (numbers per unit area for land organisms and numbers per unit volume in aquatic systems) or even weight rather than raw numbers. What may be described as an "individual" depends on the kind of organism and the aim of the scientific inquiry. In most animals, the life cycle starts with a fertilized egg, passes through a largely irreversible process of coupled growth and differentiation, and ends in a tightly integrated, unitary, adult organism. Population size can be measured by merely counting adult units and their juvenile progeny. However, in most plants and some colonial animals, growth and differentiation proceed in a modular fashion; in these cases, growth involves the replication of a basic body unit, so a fertilized egg generates a spatially distributed "population" of connected modules. In general, modular organisms show tremendous plasticity in size and form.
There are only four ways that a population can change in size: birth, death, immigration, and emigration. If immigration and emigration are negligible, the population is closed, and the difference between birth and death rates drives its dynamics. Terrestrial animals on islands often have closed populations. If immigration and emigration are important, however, the population is open, and its abundance may be substantially influenced by spatially distant events. For example, the number of barnacles that are found on a rocky coastline often reflects the density of setting larvae, which in turn is governed by events in offshore waters. If a population that is under study is found to be highly open, the spatial scale of the study may be too narrowly circumscribed to capture the important mechanisms of its population dynamics. See also: Island biogeography
Basic population models
Populations exhibit a great variety of dynamical patterns, including explosive outbreaks, local extinctions, and regular cycles or relatively constant abundances. To help describe and explain these patterns, ecologists rely on population models. Simple life cycles and closed populations provide a useful starting point in developing population models. For example, many temperate-zone insects have one annual generation; thus, at any given time, all individuals are at the same stage of life.
Iterations of discrete time-growth models for subsequent generations allow one to project population numbers through time. The theoretical framework of population ecology largely consists of elaborations of basic growth models, including extensions to more complicated life cycles and multiple species.
In addition, exponential growth has a snowballing effect—for example, the more individuals that there are, the faster that the population grows (if the growth rate is constant). Even low growth rates eventually lead to populations of enormous sizes. Some natural populations show transient phases of exponential growth, particularly in colonizing episodes. In addition, exponential growth during colonization often involves expansion across space and an increase through time. Thus, immigration and emigration, if appropriate, need to be considered.
Population heterogeneity
Population models, when interpreted literally, assume that all members of a population are identical. This is rarely true, though. Birth and death rates typically vary as a function of age, body weight, and genotype. A great deal of work in population ecology is devoted to elucidating age-specific schedules of mortality and fecundity, using these patterns to predict population growth, and interpreting these patterns in the light of evolutionary theory. To study age-structured population dynamics, the number of individuals in each age class must be monitored. The two ingredients needed to project changes in population size and age structure are the mortality schedule or survivorship curve, which describes the fraction of newborns surviving to each age, and the fecundity schedule, which describes the rate of female births per female at each age. It is a formidable task to measure complete fecundity and mortality schedules in natural populations. However, if these schedules are estimated, geometric growth models can be generalized to a matrix model. Moreover, if these schedules are constant, a population will (with rare exceptions) eventually settle into a stable age distribution in which each age class comprises a constant fraction of the total population. A population in its stable age distribution grows geometrically at a rate of increase uniquely determined from the mortality and fecundity schedules. A population displaced from its stable age distribution may exhibit transient phases of growth or decline, diverging from its long-term growth pattern.
All populations are genetically variable. If different genotypes have different fecundity or mortality schedules, genetic variation can influence population dynamics. Models that simultaneously incorporate changes in genetic composition and population growth can be quite complex. However, they may be important in describing some populations and are necessary for linking population ecology with evolutionary theory. See also: Population genetics
Population limitation and regulation
Populations cannot expand exponentially forever. Often, mortality rates increase or birth rates decrease (or both) as a result of competition for limited resources. The underlying mechanisms in these cases are called negatively density-dependent factors, which are specific examples of the more general concept of feedback. Alternatively, environmental conditions (for example, climatic shifts) can worsen, leading to population declines. Such causes for variation in birth or death rates are called density-independent factors. Ecologists have long disputed the relative importance of density-dependent and density-independent factors in determining population size. It is likely that both are important, but to differing degrees in different species and environments.
Intriguingly, population regulation by no means implies population stability. In general, a population is stable if it returns to equilibrium following a perturbation. Moreover, many local populations may not be persistent over long periods of time and thus may not be regulated in the usual sense. For example, open populations, by definition coupled by dispersal with other populations, can become reestablished by immigration following a local extinction. See also: Population dispersal
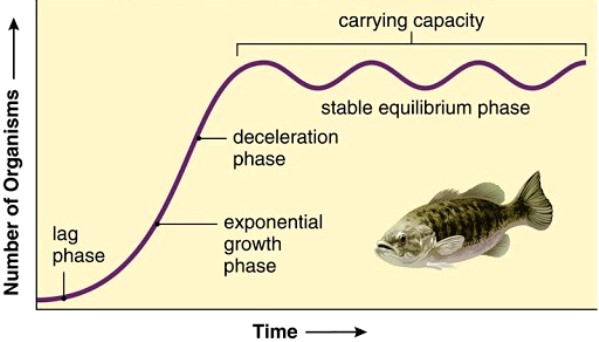
A useful method for considering the interplay of density-dependent and density-independent factors in determining population size is to plot birth and death rates as functions of density. The carrying capacity of a population in a given environment is defined to be the largest number of individuals for which the birth rate just matches the death rate (Fig. 2). In general, density-dependent factors are necessary to regulate populations, but density-independent factors must also be considered to understand fully what limits populations to a given value of carrying capacity.
Mechanisms of density dependence
Given that density dependence exists, the mechanisms generating it can be used both to predict the consequences of environmental change for population dynamics and to provide insight into systems where experimental manipulations are difficult. Density dependence often arises from competition, which is said to exist when organisms utilize common limiting resources and thereby negatively affect each other.
There are two principal sorts of competition—namely, interference and exploitative. Interference competition occurs when one individual directly harms another. Interference may be dramatic, as in lethal aggression, or subtle, as when social interactions reduce the time available for gathering resources or increase the risk of predation. Exploitative competition occurs when one individual consumes a resource, such as food, that otherwise would have been consumed by another individual. Because exploitative competition is mediated indirectly through a shared resource base, it can be more difficult to demonstrate than interference competition.
Population regulation and interspecific interactions
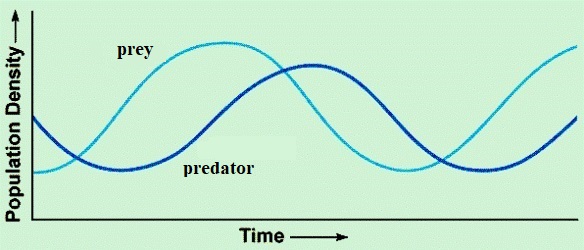
Negative density dependence may arise from interspecific interactions. A schematic classification of interactions between two species comes from considering the positive (+) or negative (−) effect that individuals of one species have on the growth rate of the other. In interspecific competition, the interaction is (−,−); in mutualism, it is (+,+). Natural enemies, defined broadly to include predators, herbivores, and parasites, are often engaged in (+,−) relations with their prey or hosts. Most species are potential prey to one or more natural enemies; even top-level carnivores may be beset by parasites.
Obviously, competitors and mutualists can dramatically affect the size of a given population and thus must be considered when studying population limitation. However, natural enemies are far more likely to be regulatory agents than are either competitors or mutualists. If two species are competing and one increases in density, the other will decrease. This will relax the interspecific competition on the first, which can then increase even more. Hence, competitive loops (and similarly mutualist loops) tend to produce positive feedback and will not regulate population growth. By contrast, predator-prey interactions (Fig. 1) may produce negative density dependence acting across several time scales on both the predator and prey. As prey in one habitat patch become more numerous, predators may almost immediately become more active or switch over from other prey types or patches. Predators also may show an intergenerational numerical response to increased prey availability. Because an increase in predator numbers usually decreases prey numbers, this induces delayed density dependence in both the predator and its prey. See also: Predator-prey interactions
Applied population ecology
In addition to its intrinsic conceptual appeal, population ecology has great practical utility. Control programs for agricultural pests or pathogens that cause human diseases ideally attempt to reduce the intrinsic rate of increase of those organisms to very low values. Analyses of the population dynamics of infectious diseases have successfully guided the development of vaccination programs. In the exploitation of renewable resources, such as in forestry or fisheries biology, population models are required in order to devise sensible harvesting strategies that maximize the sustainable yield extracted from exploited populations. Conservation biology is increasingly concerned with the consequences of habitat fragmentation for species preservation. Population models can help characterize minimum viable population sizes below which a species is vulnerable to rapid extinction, and can help guide the development of interventionist policies to save endangered species. Finally, population ecology must be an integral part of any attempt to bring the world's burgeoning human population into harmonious balance with the environment. See also: Endangered species; Fisheries ecology; Forestry

