
Key Concepts
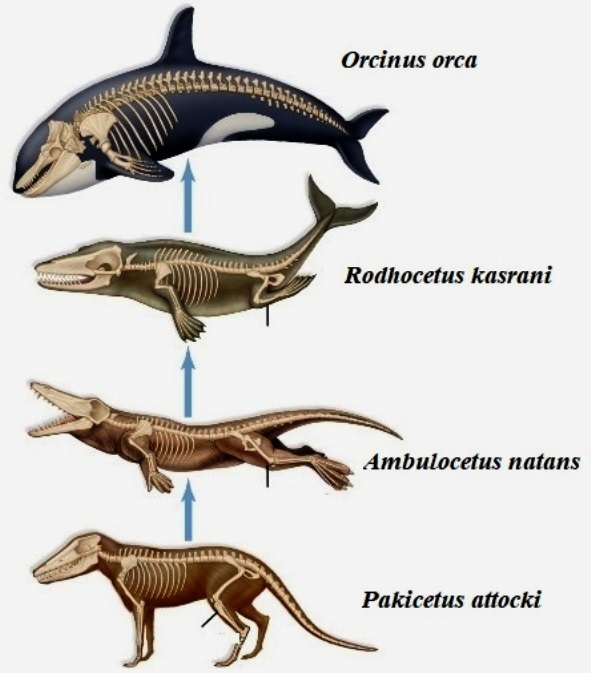
The process by which new species of organisms evolve from preexisting species. Speciation is the establishment of new, distinct species (Fig. 1). It is part of the whole process of organic evolution. The modern period of its study began with the publication of Charles Darwin's and Alfred Russell Wallace's theory of evolution by natural selection in 1858, and Darwin's On the Origin of Species in 1859. See also: Animal evolution; Charles Darwin and his theory of evolution; Evolution; Macroevolution; Plant evolution; Species concept
Belief in the fixity of species was almost universal before the middle of the nineteenth century. Then, it was gradually realized that all species continuously change, or evolve, although the causative mechanism remained to be identified. However, Darwin proposed a mechanism and postulated two points: (1) there is always some heritable variation within any species population, with the individuals differing among themselves in structure, physiology, and behavior; and (2) natural selection acts upon this variation by eliminating individuals who are less fit. Thus, if two members of an animal population differ from each other in their ability to find a mate, obtain food, escape from predators, resist the ravages of parasites and pathogens, or survive the rigors of climate, then the more successful member will be more likely than the less successful member to leave descendants. The more successful individual is said to have greater fitness, to be better adapted, or to be selectively favored. This situation applies to plants as well. For example, one plant individual is fitter than another if its heritable characteristics make it more successful than the other in obtaining light, water, and nutrients; in protecting itself from herbivores and disease organisms; or in surviving adverse climatic conditions. Over the course of time, as the fitter members of a population leave more descendants than the less fit, their characteristics become more common.
Overall, the process of natural selection tends to preserve the well adapted at the expense of the ill adapted in a variable population. The genetic variability that must exist if natural selection is to act is generated by genetic mutations in the broad sense, including chromosomal rearrangements together with point mutations. See also: Genetics; Mutation
If two separate populations of a species live in separate regions, being exposed to different environments, natural selection will cause each population to accumulate characteristics adapting it to its own environment. The two populations will thus diverge from each other and, given time, will become so different that they are no longer interfertile. At this point, speciation has occurred; that is, two species have come into existence in the place of one. This mode of speciation—speciation by splitting—is probably the most common mode. Two other modes are hybrid speciation and phyletic speciation; however, many biologists do not regard the latter as true speciation.
Splitting
There are four ways in which a species population can become separated into two (or more) parts that can undergo genetic divergence and evolve into separate species (Fig. 2). The splitting types are designated Ia, Ib, II, and III.
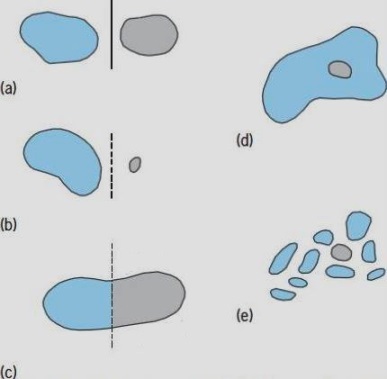
In type Ia, an extensive population becomes split into two comparatively large parts [part (a) of Fig. 2]. In terrestrial species, this may happen because movement of the Earth's crustal plates rafts the parts away from each other; alternatively, a barrier (for example, a mountain range or a tract of desert) may form and split a once-continuous population. A marine population is split if a land bridge forms that cuts through it; for example, the fall in sea level that accompanied the Pleistocene glaciations created a land bridge between Alaska and Siberia.
A type Ib split occurs if a small, outlying segment of a population becomes detached from the rest [part (b) of Fig. 2]. Such a peripheral isolate is usually a colony founded by a few emigrants from the main body of the species population. The colony can evolve into a new species even if it is not totally isolated from the parent population, provided its necessarily small gene pool chances to be well adapted to the habitat in which it finds itself. The "preadapted" possessors of these genes will be selectively favored within their small local habitat and will quickly proliferate.
Population splits of both types Ia and Ib cause the diverging populations to be geographically separated, or allopatric. The resultant genetic separation is known as allopatric speciation.
In type II, a population becomes split into geographically contiguous parts between which there is a limited amount of gene exchange. The environments of the daughter populations are assumed to differ sufficiently for natural selection to override the effects of the restricted gene flow, and speciation can thus occur. This is called parapatric speciation [part (c) of Fig. 2].
A type III split occurs if, within a single geographic region, parts of a population in some way become so isolated from each other that gene exchange between them falls below the level needed to maintain the integrity of the original population. The result is known as sympatric speciation [part (d) of Fig. 2]. It is probably common among parasites and parasitoids, but is thought to be rare or nonexistent in free-living organisms. Even though closely related species adapted to different habitats, often with different modes of life or with different resource requirements, can co-occur in the same geographic area, their co-occurrence should not be ascribed to sympatric speciation unless a mechanism can be found that could have initiated their divergence in the first place. A more likely explanation for such co-occurrences is that the species evolved allopatrically or parapatrically, that ecological divergence accompanied speciation, and that the new species then became secondarily sympatric by migrating into each other's areas after the geographic barrier that formerly separated them disappeared. See also: Island biogeography
Descriptions of allopatric, parapatric, and sympatric speciation often tacitly assume that, in the absence of clear, unmistakable barriers, gene exchange occurs freely throughout very extensive populations; therefore, an entire species is treated as a single breeding population. However, some researchers have argued that the total population of a whole species is typically broken into a large number of small breeding populations (demes) that are fairly isolated from each other and among which gene exchange is comparatively rare. Thus, a deme (rather than a species population) is the unit of evolution. As a result, sympatric speciation will sometimes seem to have occurred if a deme near the center of a species' range chances to diverge from the surrounding demes. In this case, the process may be called quasisympatric speciation [part (e) of Fig. 2]. The fact that all demes do not diverge from one another, giving as many species as there are demes, presumably results from stabilizing selection. Stabilizing selection causes groups of demes to persist in their resemblance to each other, and such groups are the "populations" constituting the visible units of evolution. See also: Population genetics
Isolating mechanisms
Two genetically diverging populations are not separate species so long as it remains possible for them to lose their distinctness, by interbreeding, if they chance to become sympatric. For speciation to be complete, the populations must evolve isolating mechanisms that prevent gene exchange between them; this allows them to maintain their genetic integrity.
There are many kinds of isolating mechanisms. The diverging populations may become genetically incompatible, so hybrids are never produced or are sterile. Even when the diverging populations can be shown to be infertile, they may be effectively isolated by a variety of mechanisms. For example, they may breed at different times or in different habitats; they may have evolved different patterns of mating behavior, different mating calls, or different sex attractants (pheromones), so individuals of the two populations never attempt to copulate; copulation may be mechanically impossible because of the structure of the genitalia (this occurs in some insects); and in insect-pollinated plants, diverging populations sometimes become adapted to different species of insect pollinators.
Hybrid speciation
In plants, new species are sometimes formed abruptly by hybrid speciation via the following mechanism. The accidental crossing of two diploid plants of different, although related, species yields a healthy (but sterile) diploid hybrid. The hybrid is sterile because the chromosomes that it has received from its parents cannot form normal bivalent pairs; meiosis (the set of two successive cell divisions that serve to separate homologous chromosome pairs and thus reduce the total number of chromosomes by half) is upset, and functional gametes cannot be produced. However, chromosome doubling occurs in the sterile hybrid, and it becomes tetraploid; normal meiosis becomes possible; fertile gametes are produced; and a new species appears. The new species is called an amphidiploid (or allotetraploid). See also: Chromosome; Meiosis
Speciation in uniparental organisms
Many plants reproduce uniparentally for long periods of time. Uniparental reproduction occurs in two ways—by autogamy (self-fertilization) and by apomixis (asexual reproduction); the latter term includes both vegetative multiplication and agamospermy (the asexual formation of viable seeds). However, a uniparental plant is not a member of a "biological species" as the term is usually defined (namely, a reproductively isolated collection of interbreeding individuals). This has led to the term "evolutionary species," a more inclusive category, which is defined so as to apply to both uniparental and biparental organisms. An evolutionary species is thus a collection of genotypically similar individuals of common ancestry whose similarity is maintained by stabilizing selection in a given environment. See also: Plant reproduction
Phyletic evolution and chronospecies
Consider a series of fossils spanning a long interval of time (long in the geological sense) and believed to represent the successive forms of a single lineage. If there has been appreciable evolution within the lineage, the change is described as phyletic evolution or anagenesis.
Fossils from a single lineage, but of different ages, may differ from each other so conspicuously that paleontologists find it convenient to treat them as taxonomically different species, even though the younger is directly descended from the older. Such species are called chronospecies, successional species, or paleospecies. They are not species in the usual sense, and questions as to whether they can interbreed do not arise because they are never alive at the same time. In a lineage evolving gradually, decisions on where to put the dividing line between each chronospecies and its successor are arbitrary. A "vanished" chronospecies, that is, one that has disappeared and been replaced by a successor chronospecies of the same lineage, is said to have undergone pseudoextinction.
Figure 3 contrasts the behavior of a lineage evolving phyletically (lineage 2) with one persisting unchanged (lineage 1). The "species" labeled B and C in lineage 2 are chronospecies. At an arbitrary point on the time scale, B becomes pseudoextinct and is succeeded by C.
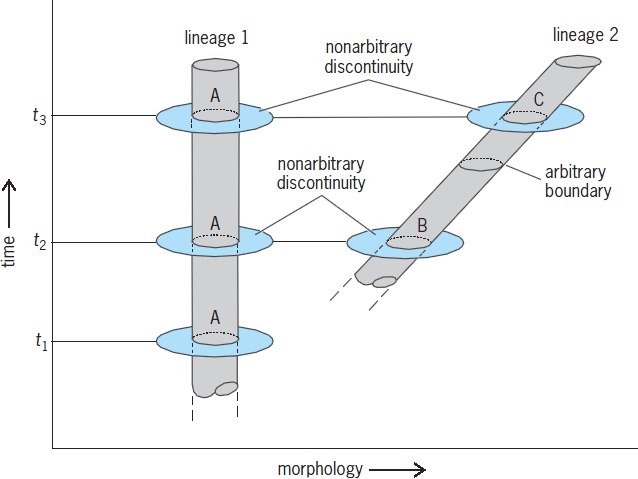
The transition from a chronospecies to its successor is not usually regarded as speciation in the strict sense. Speciation as usually defined entails the splitting of lineages.
Quantum speciation versus phyletic gradualism
When a lineage splits, the two descendant lineages may diverge slowly or rapidly. There has been much debate on whether one or the other of these modes has dominated the course of evolution. The two modes to be contrasted are shown diagrammatically in Fig. 4.
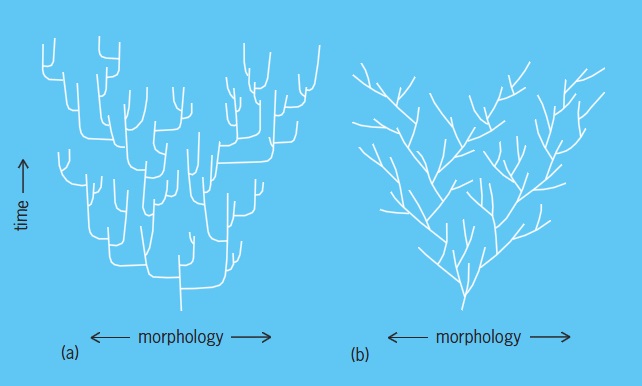
The phylogenetic tree in part (b) of Fig. 4 represents evolution and speciation according to the model known as phyletic gradualism. It supposes that all lineages undergo continuous gradual change at a fairly constant rate. When splits occur, the descendant lineages diverge gradually from each other. There is no appreciable change in evolutionary rate at the time that a split takes place. In addition, the separated lineages are still capable of interbreeding to a limited extent (although with reduced fertility), and they are known as semispecies. Their morphological divergence may sometimes be so slow that they remain practically indistinguishable even after they have lost the capacity to interbreed and have reached specific rank; if so, they are known as cryptic species.
In part (a) of Fig. 4, the phylogenetic tree represents the punctuated equilibrium model of evolution and speciation. It supposes that the splitting of a parent species population is always (or nearly always) accompanied by quantum speciation, that is, the extremely rapid (in the geological sense) and pronounced divergence of one of the descendant lineages from the ancestral form. It is assumed that the divergent species originates most often by the chance splitting off of a small peripheral isolate from an extensive parental population; this is allopatric speciation of type Ib [part (b) of Fig. 2]. If the isolate is small, its small gene pool may not be at all representative of the parent gene pool from which it is drawn. It may, simply by chance, deviate markedly. Furthermore, because of its peripheral location, the environment of the isolate is likely to differ from that of the main body of the species population, and natural selection will then tend to magnify the deviation.
According to this model, evolutionary changes associated with the appearance of new species are concentrated in speciation events of very short duration; they are represented by the horizontal segments of the branches of the phylogenetic tree [part (a) of Fig. 4]. During the long time intervals between successive speciation events, a species remains almost unchanged, as shown by the vertical segments of the branches. While a species remains in this static condition, phyletic evolution may bring about minor adjustments; this within-species evolution is known as microevolution. However, the magnitude of these phyletic changes is negligible compared with that accompanying macroevolution, or the formation of new species.
Many students of evolution believe that most groups of organisms evolve in accordance with the punctuated equilibrium model rather than by phyletic gradualism. There are two chief arguments for this view. First, it is clear from the fossil record that many species persist without perceptible change over long stretches of time and then suddenly make large quantum jumps to radically new forms. Second, phyletic gradualism seems to be too slow a process to account for the tremendous proliferation of species needed to supply the vast array of living forms that have come into existence since life first appeared on Earth.

