
Key Concepts
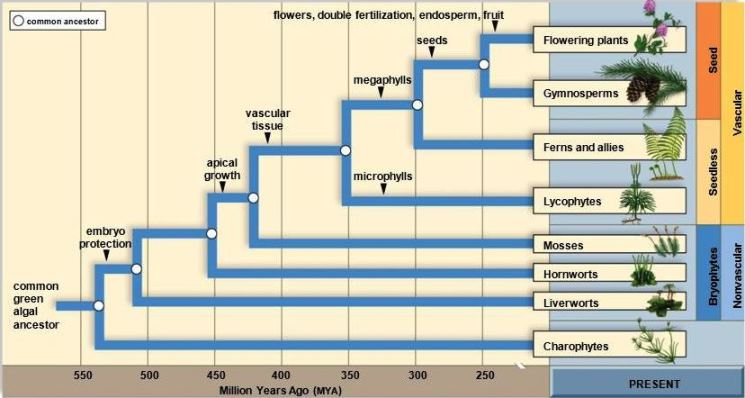
The process of biological and organic change within the plant kingdom by which the characteristics of plants differ from generation to generation. Understanding of the course of plant evolution (Fig. 1) is based on several lines of evidence. The main levels (grades) of evolution have long been clear from comparisons among living plants, but the fossil record has been critical in dating evolutionary events and revealing extinct intermediates between modern groups, which are separated from each other by great morphological gaps (evolutionary changes in many characters). Plant evolution has been clarified by cladistic methods for estimating relationships among both living and fossil groups. These methods attempt to reconstruct the branching of evolutionary lines (phylogeny) by using shared evolutionary innovations (for example, presence of a structure not found in other groups) as evidence that particular organisms are descendants of the same ancestral lineage (a monophyletic group, or clade). See also: Botany; Epigenetics and plant evolution; Fossil; Macroevolution; Organic evolution; Paleobotany; Phylogeny; Plant; Plant kingdom; Plant phylogeny; Plant taxonomy
Most botanists restrict the term plants to land plants, which invaded the land from water. There is abundant evidence of photosynthetic life extending back 3.5 billion years to the early Precambrian, notably in the form of microfossils resembling cyanobacteria (formerly referred to as prokaryotic blue-green algae) and limestone reefs (stromatolites) made by these organisms. Larger cells representing eukaryotic algae appear in the late Precambrian, followed by macroscopic algae and animals just before the Cambrian. See also: Algae; Cyanobacteria; Eukaryota; Photosynthesis; Prokaryote; Stromatolite
Origin of land plants
Cellular, biochemical, and molecular data place the land plants among the green algae, specifically the charophytes, which resemble land plants in their mode of cell division and differentiated male and female gametes (oogamy). Land plants themselves are united by a series of innovations not seen in charophytes, and many of these are key adaptations required for life on land. Charophytes have a haploid life cycle, in which the zygote undergoes meiosis (a set of two successive cell divisions that serve to separate homologous chromosome pairs and thus reduce the total number of chromosomes by half), and single-celled reproductive structures; in contrast, land plants have an alternation of generations, with a haploid, gamete-forming (gametophyte) phase and a diploid, spore-forming (sporophyte) phase. Their reproductive organs (egg-producing archegonia, sperm-producing antheridia, and spore-producing sporangia) have a protective layer of sterile cells. The sporophyte, which develops from the zygote, begins its life inside the archegonium. The spores, produced in fours by meiosis, are air-dispersed, with a resistant outer wall that prevents desiccation. See also: Cell division; Charophyceae; Meiosis; Plant reproduction
Land plants have been divided traditionally into nonvascular plants (bryophytes) and vascular plants (tracheophytes). These differ in the relative role of the sporophyte, which is subordinate and permanently attached to the gametophyte in bryophytes, but dominant and independent in vascular plants. In vascular plants, tissues are differentiated into an epidermis with a waxy cuticle that retards water loss and stomates for gas exchange, parenchyma for photosynthesis and storage, and water- and nutrient-conducting cells (xylem and phloem). However, cladistic analyses imply that some bryophytes are closer to vascular plants than others. This implies that the land-plant life cycle originated before the full suite of vegetative adaptations to life on land, and that the sporophyte began small and underwent a trend toward elaboration and tissue specialization. See also: Adaptation (biology); Bryophyta; Epidermis (plant); Parenchyma; Phloem; Plant primary vascular system; Xylem
In the fossil record, the first recognizable macroscopic remains of land plants are Middle Silurian vascular forms with a branched sporophyte; these forms are known as rhyniophytes. They differed from modern plants in having no leaves or roots; they had only dichotomously branching stems with terminal sporangia. However, spore tetrads formed by meiosis are known from older beds (Middle Ordovician); these may represent more primitive, bryophytic plants. See also: Rhyniophyta
In one of the most spectacular adaptive radiations in the history of life, vascular plants diversified through the Devonian. At the beginning of this period, vegetation was low [perhaps at most 30 cm (12 in.) in height] and probably confined to wet areas; however, by the Late Devonian, size had increased in many lines, resulting in large trees and forests with shaded understory habitats. Of the living groups of primitive vascular plants, the lycopsids (club mosses), with pointed, one-veined leaves (microphylls), branched off first, along with the extinct zosterophyllopsids, which were leafless, but resembled lycopsids in having lateral sporangia. A second line, the trimerophytes, which showed incipient differentiation of a main trunk and side branches, as well as terminal sporangia with longitudinal dehiscence, gave rise to sphenopsids (horsetails; Fig. 2), with simple leaves in a whorled arrangement, and ferns (filicopsids), with pinnately compound leaves (fronds) derived from whole branch systems. This radiation culminated in the coal swamp forests of the Late Carboniferous, with tree lycopsids (Lepidodendrales), sphenopsids (Calamites), and ferns (Marattiales). Remains of these plants make up much of the coal of Europe and eastern North America, which were then located on the Equator. See also: Coal; Coal paleobotany; Lepidodendrales; Lycopodiales; Marattiales; Sphenophyta; Tree

Seed plants
Perhaps the most significant event after the origin of land plants was evolution of the seed. Primitive seed plants (gymnosperms) differ from earlier groups in their reproduction, which is heterosporous (producing two sizes of spores), with separate male and female gametophytes packaged inside the pollen grain (microspore) and the ovule (a sporangium with one functional megaspore, surrounded by an integument, which develops into the seed), respectively. The transfer of sperm (two per pollen grain) from one sporophyte to another through the air, rather than by swimming between gametophytes living on or in the soil, represents a step toward independence from water for reproduction, and it must have helped plants invade drier areas than they had previously occupied. In addition, seed plants have new vegetative features, particularly secondary growth, which allows production of a thick trunk made up of secondary xylem (wood) surrounded by secondary phloem and periderm (bark). Together, these innovations have made seed plants into the dominant organisms in most terrestrial ecosystems ever since the disappearance of the Carboniferous coal swamps. See also: Palynology; Periderm; Pollen; Seed
A major breakthrough in understanding the origin of seed plants was recognition of the progymnosperms (for example, Archaeopteris) in the Middle and Late Devonian. These plants, which were the first forest-forming trees, had secondary xylem, phloem, and periderm, but they still reproduced by spores, implying that the anatomical advances of seed plants arose before the seed. Similar to sphenopsids and ferns, they were apparently derived from trimerophytes. The earliest seed plants of the Late Devonian and Carboniferous, called seed ferns because of their frondlike leaves (a convergence with true ferns), show steps in the origin of the seed, including the fusion of branchlets surrounding the megasporangium into the integument. Origin of the typical mode of branching in seed plants, from buds in the axils of the leaves, occurred at about the same time. See also: Pteridosperms
Among seed plants, coniferopsids (fossil cordaites, living conifers, and possibly ginkgos), with fan-shaped to needlelike leaves, have often been considered an independent line of evolution from progymnosperms. However, cladistic analyses indicate that coniferopsids were derived from seed ferns with coniferlike pollen and seeds. This event may have involved the elimination of fronds and their replacement by scale leaves, which occur around the buds in all seed plants. Cordaites and conifers both appear in the Late Carboniferous; comparisons of their early members indicate that the seed-bearing cone of conifers is a complex structure, in which the cone scales are actually modified branches. See also: Cordaitales; Pinales
Seed plants became dominant in the Permian during a shift to drier climates and following extinction of the coal swamp flora in the European-American tropical belt. Early conifers, with small, needlelike leaves, predominated in the tropics, and extinct glossopterids, with simple, deciduous, net-veined leaves, inhabited Gondwana (the ancient continent that fragmented and drifted apart during the Triassic to form eventually the present continents). Climate moderation in the Triassic coincided with the appearance of new seed plant groups, including the living cycads and ginkgos (today reduced to one species, Ginkgo biloba) and the extinct Bennettitales (Cycadeoidales), as well as more modern ferns. Many Mesozoic groups show adaptations for protection of seeds against animal predation, whereas flowers of the Bennettitales constitute the first evidence for attraction of insects for cross-pollination. See also: Coevolution between flowering plants and insect pollinators; Cycadeoidales; Pollination
Angiosperms
The last major event in plant evolution was the origin of angiosperms (flowering plants; Magnoliophyta). Angiosperms show numerous innovations that allow more rapid and efficient reproduction; hence, they are the seed plants that dominate the modern flora. The flower, typically made up of protective sepals, attractive petals, pollen-producing stamens, and ovule-producing carpels (all considered modified leaves), favors more efficient pollen transfer by insects. The ovules are enclosed in the carpel, so the pollen germinates on the sticky stigma of the carpel rather than in the pollen chamber of the ovule. The carpels (separate or fused) develop into fruits, which often show special adaptations for seed dispersal. Other advances include an extreme reduction of the gametophytes, as well as double fertilization, whereby one sperm fuses with the egg, and the second sperm fuses with two other gametophyte nuclei to produce a triploid, nourishing tissue called the endosperm. Angiosperms also developed improved vegetative features, such as more efficient water-conducting vessels in the wood and leaves with several orders of reticulate venation. See also: Angiosperm phylogenetics; Angiosperms; Flower; Fossil seeds and fruits; Magnoliophyta; Origin of the flowering plants
Most botanists believe that the most primitive living angiosperms are magnoliid dicots, based on their gymnospermlike pollen, wood anatomy, and flower structure. Studies of Cretaceous fossil pollen, fossil leaves (Fig. 3), and fossil flowers confirm this view by showing a rapid (but orderly) radiation beginning with magnoliidlike and monocotlike types, followed by primitive eudicots, including some related to sycamores and lotuses. See also: Dicotyledons; Eudicotyledons; Monocotyledons

Morphological and molecular data imply that angiosperms are monophyletic and most closely related to Bennettitales and Gnetales (a seed plant group that also radiated in the Early Cretaceous). Because all three groups have flowerlike structures, suggesting that the flower and insect pollination arose before the closed carpel, they have been called anthophytes. These relationships, plus problematical Triassic pollen grains and macrofossils with a mixture of angiospermlike and more primitive features, suggest that the angiosperm line goes back to the Triassic, although perhaps not as fully developed angiosperms. Within angiosperms, it is believed that magnoliids are relatively primitive, monocots and eudicots are derived clades, and wind-pollinated temperate trees (for example, oaks, birches, and walnuts) are advanced eudicots. However, magnoliids include both woody plants (for example, magnolias and laurels) and herbs (for example, water lilies and peppers), and their flowers range from large, complex, and insect-pollinated to minute, simple, and wind-pollinated. These extremes are present among the earliest Cretaceous angiosperms, and cladistic analyses disagree on which is most primitive. See also: Gnetales
Although plant extinctions at the end of the Cretaceous have been linked with the radiation of deciduous trees and the proliferation of fruits dispersed by mammals and birds, these extinctions were less dramatic than those in the animal kingdom. During the Early Tertiary, climates were still much milder than today, but mid-Tertiary cooling led to contraction of the tropical belt and expansion of seasonal temperate and arid zones. These changes led to the diversification of herbaceous angiosperms, such as composites and grasses, and the origin of open grassland vegetation, which stimulated the radiation of hoofed mammals and ultimately the invention of human agriculture. See also: Agriculture; Extinction; Fruit; Origin of grasslands

