
Key Concepts
The transport of pollen grains from the plant parts that produce them to the ovule-bearing organs, or to the ovules (seed precursors) themselves. Pollination is a process that ensures the successful life cycle of plants. Without pollination, there would be no plant fertilization; thus, it is of crucial importance for the production of fruit crops and seed crops. In the pollination process, pollen (the small male reproductive bodies produced in the pollen sacs of seed plants; Fig. 1) is transferred to the female reproductive organs of a plant, thus facilitating fertilization. In gymnosperms (plants with naked seeds), including conifers and cycads, the pollen, usually dispersed by the wind, is simply caught by a drop of fluid excreted by each freely exposed ovule. In angiosperms (flowering plants), where the ovules are contained in the pistil, the pollen is deposited on the pistil's receptive end (the stigma), where it germinates. Pollination also plays an important part in plant-breeding experiments that are aimed at increasing crop production through the creation of genetically superior types. See also: Agricultural science (plant); Fertilization (plant); Flower; Palynology; Plant breeding; Plant reproduction; Pollen; Seed; Seed germination
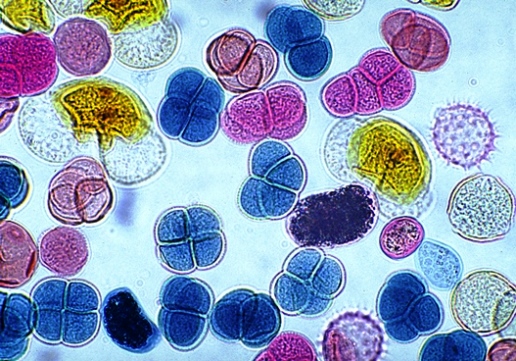
Ideally, each pollen grain forms a pollen tube, which grows down through the slender style (stalk of the pistil) toward one of the ovules enclosed in the basal part or ovary. Here, one of the two sperm cells within the pollen tube fuses with the egg cell of the ovule, making possible the development of an embryo, whereas the other sperm cell combines with the two polar nuclei of the ovule, thus starting the formation of endosperm (a tissue rich in reserve food). After this double fertilization, the ovule enlarges and transforms into a seed. See also: Plant anatomy
Self- and cross-pollination
In most plants, self-pollination is difficult or impossible. For example, in dichogamous flowers, the pistils and stamens reach maturity at different times: in protogyny, the pistils mature first; and in protandry, the stamens mature before the pistils. Selfing is also impossible in dioecious species, such as date palms and willows, where some plants bear flowers that have only pistils (pistillate or female flowers), whereas other individuals have flowers that produce only pollen (staminate or male flowers). In monoecious species, such as hazel, where pistillate and staminate flowers are found on the same plant, self-breeding is at least reduced. Heterostyly, as exemplified by certain primroses, is another device that promotes outbreeding (the union of gametes from organisms that are not closely related). Here, some flowers (pins) possess a long pistil and short stamens, whereas others (thrums) exhibit the reverse condition; each plant individual bears only pins or only thrums. Pollen transport from stamens to pistils of the same length is easy; however, in the other cases, it is difficult.
Flower attractants
As immobile organisms, plants normally need external agents for pollen transport. These agents can be insects (Fig. 2), wind, birds, mammals, or water, roughly in that order of importance. For example, in certain arum lilies, Dutchman's pipes, orchids, and other plants, the pollinators are simply trapped. In the large majority of cases, though, the flowers offer one or more rewards to pollinators, such as sugary nectar, oil, solid food bodies, perfume, sex, an opportunity to breed (as in figs), a place to sleep (Serapias orchids), or some of the pollen itself. For the attraction of pollinators, flowers provide either visual or olfactory signals. Color includes ultraviolet, which is perceived as a color by most insects and at least some hummingbird species. Fragrance is characteristic of flowers that are pollinated by bees, butterflies, and hawkmoths, whereas carrion or dung odors are produced by flowers that cater to certain beetles and flies. By using a combination of olfactory and visual signals, a few orchids (including Ophrys and Cryptostylis species) mimic females of certain bees or wasps so successfully that the corresponding male insects will try to mate with them, thus achieving pollination (pseudocopulation).

At close range, pollinating insects are guided to the nectar in many cases by special color patterns, lines, or dots, known as nectar or honey guides. Olfactory honey guides and osmophores (particular flower parts specialized for odor production) may be equally important. For better evaporation of odors, a number of flowers may also release considerable heat through a special process known as thermogenic respiration (this occurs in certain arum lilies and water lilies).
Animal pollinators
Although some flowers are generalists, catering to a whole array of different animals, others (for example, Ophrys) are highly specialized, being pollinated by only a single species of insect. Extreme pollinator specificity is an important factor in maintaining the purity of plant species in the field, even in those cases where hybridization can easily be achieved artificially in a greenhouse or laboratory, as in most orchids. The almost incredible mutual adaptation between pollinating animal and flower that can frequently be observed exemplifies the idea of coevolution (Fig. 2). See also: Adaptation (biology); Coevolution between flowering plants and insect pollinators; Mutualism; Plant-animal interactions; Pollination mutualisms by insects before the evolution of flowers
Bird pollination
Hummingbirds have good red vision and feed on the wing. Flowers pollinated by these birds are often orange and red, and they hang down freely or are exposed (Fuchsia); in addition, they are sturdy, lack a landing platform, are open in the daytime, have little or no odor, and offer a not-too-concentrated nectar. Plants with flowers pollinated by birds other than hummingbirds (such as sunbirds in Africa) usually provide perches. See also: Apodiformes
Bat pollination
Flowers pollinated by bats include those of the sausage tree, saguaro cactus, cup-and-saucer vine, and certain bananas. They are open in the evening, when they emit a rank or musty odor; their petal colors are either murky and nondescript or snow white; they are wide-mouthed and often bell-shaped; and they normally offer both nectar and pollen in abundance. Perhaps because pollinating bats have a poorly developed sonar sense, these flowers often hang down freely from ropelike twigs (flagelliflory) or grow on the main stem or limbs of the tree (cauliflory), which makes them easier to locate. However, it is also possible that these characteristics evolved to facilitate the dispersal of the fruits and seeds by bats, and that the pollinators may have been different animals. See also: Chiroptera
Hawkmoth pollination
Flowers pollinated by hawkmoths have a great deal in common with those pollinated by hummingbirds, but they are white or light-colored and very fragrant, especially in the evening when they are open.
Butterfly pollination
Flowers pollinated by butterflies open in the daytime, are usually colorful and fragrant [butterfly bush (Buddleia) and lilacs], and either offer a flat landing space (Impatiens) or, when small, are combined in inflorescences that provide flattish surfaces to be walked on (red valerian and Buddleia). The narrow flower tubes allow entry of the slender butterfly proboscis, but discriminate against other insects. See also: Lepidoptera
Fly and beetle pollination
Both flies and beetles are highly diversified groups, making it difficult to describe “typical” fly- or beetle-pollinated flowers. Trap flowers exploit types such as carrion and dung beetles (or flies), which are not at all adapted to flowers. However, many modern flowers cater to animals with highly specialized mouthparts, including certain longhorn beetles and the bee-fly Bombylius. See also: Coleoptera; Diptera
Bee pollination
The social bees, especially honeybees (Apis; Fig. 3) and bumblebees (Bombus), are the most important of all insect pollinators. Whereas honeybees are subtropical in origin, bumblebees are adapted to cold climates. The larvae of both Apis and Bombus live on pollen and honey (a modified nectar); the adults drink nectar. Among the physical and behavioral adaptations that the adults possess to obtain these foods are long tongues, hairy bodies, special pollen baskets on the hind legs, and a good color sense. To complement these attributes, the flowers pollinated by bees generally have bright colors, minty odors, and a landing platform, and their pollen grains are sticky, spiny, or highly sculptured for better adherence to the bees. In addition, honeybees have an excellent communication system: a worker bee can indicate both the direction and the distance of a rich food source by means of special dances, so that the resource can be exploited cooperatively. See also: Bee dance language; Beekeeping; Hymenoptera; Social insects

Wind pollination
Wind pollination is prevalent in the primitive cycads and in conifers, but it also is found in such familiar flowering plants as grasses, rushes, sedges, cattails, plantains, nettles, hemp, sorrel, alder, birch, hazel, poplars, and temperate-region oaks. It offers advantages in regions where animal pollinators are scarce or where they cannot operate as a result of adverse environmental or seasonal conditions, such as strong winds. Wind-pollinated flowers are usually inconspicuous, being devoid of attractants and rewards for pollinators. They lack showy petals, fragrance, and nectar. To facilitate exposure to the wind, blooming often takes place in early spring before the leaves have emerged, or the flowers may be placed very high on the plant. In a number of species, the anthers burst open in an explosive fashion, scattering the pollen widely; the stigmas often are long and divided into arms or lobes, or they are feathery (like those of some willows and cereals), thereby facilitating pollen catching. See also: Wind
Water pollination
Although the pollen grains of some plants can be made to germinate in aqueous sugar solutions, water alone in most cases will destroy them. Accordingly, water is seldom the true medium of pollen dispersal, even in aquatic plants. In ribbon weed (Vallisneria), the male and female flowers are released to the surface separately. At the surface, male flowers are driven by the wind until they bump into female flowers; the shock causes the pollen to be catapulted over to the stigma. True water dispersal of pollen (hydrophily), in which the grains are wetted, is found only in the freshwater hornworts (Ceratophyllum) and marine seagrasses (Zostera and Phyllospadix). See also: Water
Ecology
Traditionally, pollination biology has considered mechanisms that promote or guarantee the proper transfer of pollen from one individual of a given plant species to another. Often this involves a careful description of the transfer mechanism coupled to a particular pollinating agent (such as hawkmoths, bumblebees, bats, or wind). Pollinator-plant interactions, however, have also been used as a model for understanding broader evolutionary and ecological issues that pertain to all living systems. Pollination systems have proven to be an excellent model for studying many issues in population biology. In addition, because plants have a variety of breeding structures, they provide an opportunity for assessing the reproductive value of different sexual strategies and for modeling the evolution of breeding systems. Moreover, if pollen movement is considered as a measurable analog of gene movement, then pollination systems are ideal for unraveling patterns of gene flow and the concomitant genetic structuring of plant populations. Overall, the evolutionary ecology of many plants and pollinators may be best understood as a result of complex interactions among populations of many species. See also: Ecological communities; Ecological competition; Ecology; Population ecology; Population genetics

