
Key Concepts
The study of the structure of feeding relationships among organisms in an ecosystem. Trophic ecology investigates how various organisms at different feeding (trophic) levels interact within an ecosystem. In general, feeding or trophic relationships are represented as a food web or as a food chain (often depicted as a pyramid; Fig. 1). Food webs depict trophic links between all species sampled in a habitat, whereas food chains simplify this complexity into linear arrays of interactions among trophic levels. In other words, trophic levels (for example, plants, herbivores, detritivores, and carnivores) are amalgamations of species that have similar feeding habits. (However, not all species consume prey on a single trophic level; for example, omnivores are species that feed on more than one trophic level.) Overall, trophic ecologists focus on the interplay between feeding relationships and ecosystem attributes, such as nutrient cycling, physical disturbance, or the rate of tissue production by plants and the accrual of detritus (dead organic material). See also: Ecology; Ecosystem; Food web
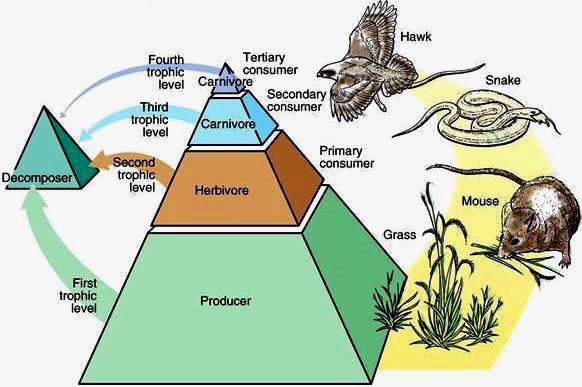
The three fundamental questions in the field of trophic ecology are as follows: (1) What is the relationship between the length of food chains and plant biomass (the total amount of plants at the bottom of the food chain)? (2) How do resource supply to producers (plants) and resource demand by predators determine the relative abundance of organisms at each trophic level in a food chain? (3) How long are real food chains, and what factors limit food chain length?
Effect of food chain length on plant biomass
A central theory in ecology is that "the world is green" because carnivores prevent herbivores from grazing green plant biomass to very low levels. Trophic structure (the number of trophic levels) determines trophic dynamics (as measured by the impact of herbivores on the abundance of plants). Indirect control of plant biomass by a top predator is called a trophic cascade. Cascades have been demonstrated to varying degrees in a wide variety of systems, including lakes, streams, subtidal kelp forests, coastal shrub habitats, old fields, grassland savannas, tundra, shrublands, and belowground soil communities. In these systems, the removal of a top predator has been shown to precipitate dramatic reductions in the abundance (or biomass) of species at lower trophic levels. Food chain theory predicts a green world when food chains have odd numbers of trophic levels; however, a barren world (plants suppressed by herbivores) is predicted in systems with even numbers of trophic levels. See also: Predator-prey interactions
The reduction and subsequent return of sea otters in coastal ecosystems provides a lucid example of cascading trophic effects in marine food chains and alternation between plant-dominated and plant-depleted subtidal habitats. Sea otters once were abundant in coastal regions from northern Japan to central Baja California, Mexico, but were reduced to a number of widely scattered remnant populations by hunting. The reduction of sea otters likely resulted in dramatic changes in prey population dynamics in ecosystems previously occupied by sea otters. For example, in rocky subtidal habitats, the abundance of sea urchins (a preferred prey item of otters) was much higher in habitats with depleted otter populations. In otter-free habitats (two trophic levels), urchin abundance was high enough to overgraze large benthic algae, including kelps. By contrast, in areas supporting remnant populations of otters (three trophic levels), urchin populations were limited to small individuals and were often restricted to cryptic habitats. As a result, kelp biomass was much higher in these areas. Therefore, odd and even food chains lead to green and barren subtidal worlds, respectively (Fig. 2). The result of the presence of otters in nearshore habitats is an increased abundance of kelps and other macroalgae, which, in turn, provide habitat and food for a number of associated species. Otters thus play a critical role in structuring subtidal communities. Otters and other species whose effects on the abundance of other species in a food chain or food web are large compared with their relative abundance in the food web are called keystone species.
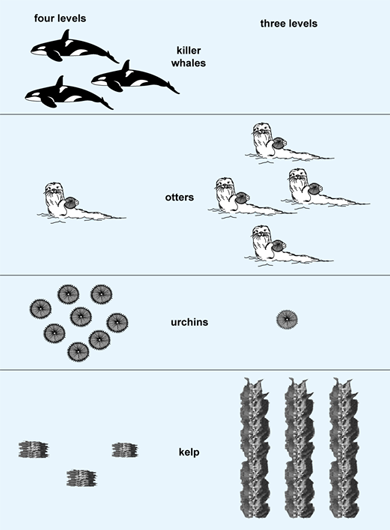
In addition, sea otters have declined in abundance in certain marine areas. A convincing hypothesis for the cause of this decline is a shift in the foraging behavior of killer whales from the once abundant baleen whales and pinnipeds (seals, sea lions, and walruses) to sea otters (Fig. 2). Concomitantly, comparison of urchin abundance, grazing rates, and kelp biomass during time periods before and after observed increases in killer whale predation on otters has revealed cascading effects of killer whales in this system. Prior to increased predation by killer whales, coastal subtidal habitats were characterized by low urchin and high kelp abundance. By contrast, these same habitats had much higher urchin densities and more barren substrates immediately following shifts in killer whale foraging and concomitant otter declines. The addition of killer whales (four trophic levels) led to a shift from green to barren worlds as a result of the otters' diminished control of urchin grazing. See also: Marine ecology
Supply and demand in food chain dynamics
Although predators often have strong indirect effects on plant biomass as a result of trophic cascades, both predation (a top-down force) and resource supply to producers (a bottom-up force) play strong roles in the regulation of plant biomass. The supply of inorganic nutrients (such as nitrogen and phosphorus) at the bottom of a food chain is an important determinant of the rate at which the plant trophic level produces tissue (primary production, or productivity) and, in some cases, of the total biomass of this trophic level. However, the degree to which nutrient supply enhances plant biomass accrual depends on two factors: (1) how many herbivores are present (which, in turn, depends on the number of trophic levels in the system) and (2) the degree to which herbivores can respond to increases in plant productivity and control plant biomass. See also: Biological productivity; Biomass
The relative importance of top-down (demand) versus bottom-up (supply) forces is well illustrated by lake systems, in which the supply of phosphorus (bottom-up force) and the presence of piscivorous (fish-eating) fish (top-down force) have significant effects on the standing stock of phytoplankton. For example, in small lakes of the Canadian Shield of North America, phytoplankton production is strongly dependent on phosphorus. In a classic experiment, a single lake was divided in half with a plastic barrier. Phosphorus, nitrogen, and carbon were added to one side of the lake, whereas just nitrogen and carbon were added to the other (control) side. Blooms of algae turned the water green on the side to which phosphorus was added, but not on the control side. This result was instrumental in convincing local and national governments in Canada and the United States to regulate phosphate release in sewage to prevent noxious, oxygen-depleting blooms of algae in lakes and other sources of drinking water. See also: Algae; Freshwater ecosystem; Lake; Phytoplankton; Zooplankton
Herbivores may be capable of counteracting nutrient-driven algal blooms, especially in relatively nutrient-poor (oligotrophic) lakes. In these lakes, the effects of top predators, such as largemouth bass, on phytoplankton are analogous to those of killer whales on kelp, and the response of phytoplankton to nutrient loading depends on the number of fish trophic levels in the system. Small lakes have up to two distinct trophic levels of fish species—those that eat zooplankton (zooplanktivores) and those that eat other fish (piscivores). In oligotrophic lakes with only zooplanktivores (that is, which have three trophic levels), these fish deplete zooplankton that would otherwise graze on phytoplankton, thereby allowing phytoplankton biomass to increase with nutrient (phosphorus) loading from the bottom of the food chain. By contrast, if piscivores, such as largemouth bass, are added to these same lakes (which then have four-trophic levels), they eat the zooplanktivores, zooplanktivore abundance declines, and zooplankton organisms recover and graze the once green lake until it is barren. In these four-trophic-level systems, phosphorus additions may increase the productivity, but not the biomass, of the plant trophic level. Thus, top-down control of phytoplankton biomass by predators is possible in oligotrophic lakes, depending on food chain length.
In more fertile (mesotrophic) lakes, piscivores may control zooplanktivore abundance, but the zooplankton organisms (herbivores) are still not capable of keeping pace with increasing phytoplankton production across gradients of increasing nutrient loading. Top-down control attenuates between the third (zooplanktivore) and second (herbivore) trophic levels, and nutrient loading has positive effects on plant productivity and biomass.
In lakes with moderate-to-high nutrient loading, high productivity by species at all trophic levels may preclude control by consumers at the top. In this case, increased nutrient levels lead to increases in the biomass of phytoplankton, zooplankton, and fish. Thus, although plant productivity may increase fairly predictably with nutrient loading, the biomass of the plant trophic level depends on both nutrient supply and trophic structure. Top-down control of plant biomass by top predators appears to be more important in nutrient-poor systems. As nutrient supply increases, the relative influence of bottom-up control on plant biomass is increasingly important.
Determinants of food chain length
There are three major hypotheses for what determines the length of food chains in nature, based on energy, resilience, and ecosystem size.
Productivity and efficiency
From an energetic perspective, food chain length is limited by two factors: the total rate of plant or bacterial production, and the efficiency at which members of each trophic level assimilate this energy as it moves up the food chain. Productivity determines the total energy supply for a system; however, not all of this energy is incorporated by successively higher trophic levels. Inefficiency in transfer, in either consumption or assimilation, reduces the fraction of total available energy propagated between each trophic level. Typical transfer efficiencies range from 5% to 15% for herbivores and carnivores (for example, zooplankton and bass). In three transfers, the total energy base of the food chain is reduced by more than 99%, limiting the energy available to an additional trophic level. Thus, given fixed transfer efficiencies, additional trophic levels may be added only with increases in productivity at the base of the food chain. Several large-scale syntheses (on lake and tundra island systems) suggest that food chain length does increase with plant productivity. Nevertheless, long food chains (more than five trophic levels) are extremely rare in nature. See also: Ecological energetics
Resilience
The idea that the upper limit to food chain length may be determined by the inherent instability of long chains derives from theoretical studies of food chains. In these studies, theoretical ecosystems are constructed as coupled differential equations describing the population dynamics of single-species trophic levels. To analyze food chain stability, disturbance is introduced by changing (reducing or increasing) the abundance at one trophic level. Resilience, an ecological metric for stability, is measured as the inverse of the time required for all trophic levels to return to their previous abundance levels after a disturbance. Longer food chains have consistently longer return times (lower resilience) than short chains (four or less trophic levels), suggesting that longer chains should be rare in nature because return times may be longer than the recurrence interval of disturbance. Although results from model food chains are consistent with field tests in small aquatic systems, the empirical mechanisms behind decreased stability of longer food chains at larger spatial scales are not as clearly developed as the logic of the productivity hypothesis. Moreover, experiments in rivers suggest that disturbance in some cases may act to lengthen food chains, suggesting that the effects of disturbance on food chain length may vary between ecosystem types. See also: Systems ecology; Theoretical ecology
Ecosystem size
One final factor that may set constraints on food chain length is ecosystem size. Size can be defined relatively easily in habitats with discrete boundaries (for example, lakes and oceanic islands). It has been hypothesized that ecosystem size and productivity would interact to determine food chain length. This theory holds that the coverage (in area or volume) of a given level of productivity would provide a more comprehensive measure of the energy supply at the base of food chains and, thus, their potential length. Food chains should increase in length with increasing "productive space" rather than with increasing productivity alone. However, the size of ecosystems alone may provide an equally robust prediction of food chain length. Both body size and home range size may increase with trophic position. Small habitats are simply not large enough to support the home range or provide ample habitat for larger carnivorous species and thus may limit the length of food chains. In lake systems, for instance, ecosystem size alone predicts more variability in food chain length than either productivity or productive space. In these systems, large piscivores are often found only in deep waters, which are habitats found only in larger lakes.
Bioaccumulation of contaminants in top predators
One application of trophic ecology has been the recognition and prevention of bioaccumulation of pesticides and heavy metals in top predators. An example of bioaccumulation is the near-demise of birds of prey, such as the bald eagle, osprey, and peregrine falcon, as a result of the formerly unrestricted application of dichlorodiphenyltrichloroethane (DDT) and other organochlorine pesticides to crop fields, lakes, and ponds to kill insect pests (crop herbivores and mosquitoes). Although DDT is not lethal to animals at higher trophic levels, it is not excreted by them. Thus, birds and fish that consume large quantities of insects or other herbivores exposed to DDT concentrate the pesticide in their tissues. Birds of prey, in turn, further concentrate DDT because they rely heavily on fish or other birds as prey species (two trophic transfers of DDT). Birds with high pesticide levels tend to lay eggs with abnormally thin shells that crack during incubation, impairing reproduction and causing dramatic population declines. Recognition of this problem led to a ban on the use of DDT and other stable pesticides in many industrialized countries. See also: Insecticide; Pesticide
Many fish species common in markets around the world are top predators and may be reservoirs for other common toxins, such as mercury. Interestingly, fish from ecosystems with long food chains may pass along more mercury to humans than those from food chains with fewer trophic levels and, thus, fewer trophic transfers of this toxin. See also: Mercury (element); Toxicology; Toxin

