Key Concepts
The set of structures, such as the heart (or pumping mechanism) and the arteries, veins, and capillaries, which provide channels for the flow of blood. The cardiovascular system (Fig. 1) is sometimes called the blood-vascular system. The circulatory system includes both the cardiovascular and lymphatic systems; the latter consists of lymph channels (lymphatics), nodes, and fluid lymph, which finally empties into the bloodstream. With regard to vertebrates, all animals in this grouping feature a closed system of branched vessels, a ventral heart, and a basic pattern of organization that ranges from the single system in most fishes to the double system in terrestrial forms. Embryologically, the cardiovascular system in vertebrates arises from the splanchnic mesoderm, with the first blood and vessels formed in the wall of the yolk sac. See also: Blood; Blood vessel; Circulation; Developmental biology; Embryology; Lymphatic system; Vertebrata; Yolk sac
Heart
The hearts of vertebrates differ from those of animals in the lower phyla in their ventral rather than dorsal location. Blood is pumped anteriorly through arteries and forced to the dorsal side. The greater part then courses posteriorly through arteries, which terminate in capillaries in various parts of the body. The blood returns to the heart through veins. The heart is usually described as tubular during its early morphological differentiation, although it is saccular in shape in most species. The morphological changes that occur in the heart while it differentiates from a tubular structure to its definitive adult form vary for different classes and species of vertebrates in correlation with different functional requirements. Complete division of the heart into right and left sides, that is, a complete separation of respiratory and systemic circulations, is achieved only in birds and mammals. See also: Aves; Heart (invertebrate); Heart (vertebrate); Mammalia
Phylogeny
In lower vertebrates, the heart is located far forward in the body, but there is a gradual backward shifting as the vertebrate scale is ascended. The heart lies in a pericardial cavity that is surrounded by an investing membrane, called the pericardium. The pericardial cavity is a portion of the coelom, which has been separated from the remainder of the body cavity. In elasmobranchs, the separation is incomplete, and the two portions of the coelom are connected by a pericardio-peritoneal canal. A thin serous membrane, the epicardium, covers the surface of the heart. It is continuous with the lining of the pericardial cavity.
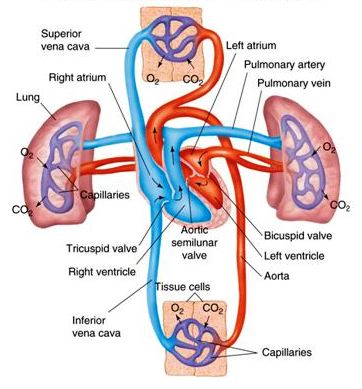
The heart is primarily a pulsating tube, and the lining (endocardium) is derived from fusion of two vitelline veins. Its muscular wall (myocardium) and the epicardium originate from the surrounding splanchnic mesoderm. It becomes divided into chambers called atria and ventricles. In addition, two accessory chambers, called the sinus venosus and conus arteriosus, may be present. Cyclostomes and most fishes have two-chambered hearts with one atrium and one ventricle. A sinus venosus connects with the atrium and a conus arteriosus leads from the ventricle. Dipnoans and amphibians have three-chambered hearts with two atria and one ventricle. The three-chambered heart of most reptiles is similar to that of amphibians, but an incomplete partition appears in the ventricle. In crocodiles and alligators, this becomes complete, forming a four-chambered heart with two atria and two ventricles. Birds and mammals have four-chambered hearts (Fig. 2). Valves are present to regulate the direction of blood flow.
Circulation in the heart
Vertebrates with two-chambered hearts have the single type of circulation (Fig. 3) in which only unoxygenated blood passes through the heart, which pumps it to the gills for aeration. The double type of circulation (Fig. 4) exists in three- and four-chambered hearts through which pass two streams of blood—one oxygenated and the other unoxygenated or partly oxygenated. Even in three-chambered hearts, these streams do not mix to any appreciable extent. Partitioning of the heart has been associated in evolution with development of pulmonary circulation accompanying the appearance of lungs. See also: Lung
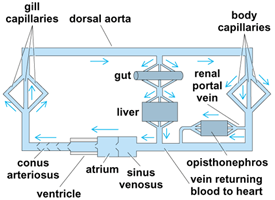
In those forms having two-chambered hearts, unoxygenated blood from all parts of the body is collected by the sinus venosus, which joins the atrium. In the three-chambered hearts of dipnoans and amphibians, the sinus venosus has shifted its position and joins the right atrium. A large sinus venosus is present in certain reptiles; in most reptiles, though, it is very small or is lacking. Birds and mammals possess a sinus venosus only during early embryonic development. In vertebrates having three- or four-chambered hearts, unoxygenated blood is returned to the heart through the sinus venosus or directly to the right atrium. In addition, the conus arteriosus no longer exists as such in adults of higher forms; it is split into trunks leading to the aorta and lungs, respectively.
The cardiac rhythm is myogenic (generated by muscles) in all vertebrates, and the heartbeat is initiated in a specialized group of muscle cells that form the pacemaker. From the pacemaker, a wave of electrical excitation passes across the heart, activating the contractile process in cardiac muscle. See also: Muscular system
Arterial system
Although the arterial systems of various adult vertebrates appear to be very different in arrangement, a study of development reveals that all systems are built upon the same fundamental plan. The arterial system consists of ventral and dorsal aortas, aortic arches, somatic arteries, and visceral arteries.
Various paired and unpaired arteries arise along the length of the dorsal aorta to supply all structures of the body posterior to the pharyngeal region. Anterior continuations of the ventral aorta and the anterior remnants of the paired dorsal aortas supply the head and anterior branchial regions. Although the vessels arising from the dorsal aorta are fairly uniform throughout the vertebrate series, the aortic arches undergo profound modifications. The changes are similar in members of a given class. Blood, pumped anteriorly by the heart, passes to the aortic arches. These vessels then carry the blood to the dorsal aorta or aortas, from which it goes either anteriorly to the head or posteriorly to the remainder of the body. Moreover, coronary arteries, which supply the tissues of the heart itself, arise as branches of certain aortic arches or of the systemic trunk near the point where it emerges from the heart.
The arteries supplying the body proper are usually paired structures that clearly show evidences of segmental arrangement (metamerism). They spring from the dorsolateral regions of the dorsal aorta. Fusion of two or more segmental arteries may take place, obscuring the fundamental metameric arrangement.
Arteries supplying the viscera are of two kinds—paired and unpaired. The paired arteries, usually restricted to certain regions, are segmentally arranged. They supply structures derived from the part of the embryo from which the urogenital organs and their ducts arise. Unpaired visceral arteries supply the spleen and digestive tract. There are usually three such vessels in vertebrates. The celiac artery supplies the stomach, spleen, pancreas, liver, and duodenum; the superior mesenteric artery supplies most of the intestine and a portion of the pancreas; and the inferior mesenteric artery goes to the posterior part of the large intestine and rectum. Variations from this condition are accounted for by fusions or separations.
Venous system
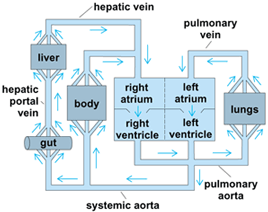
A comparison of veins in the various vertebrate groups shows that they too are arranged according to the same fundamental plan and that the variations encountered form a logical sequence as the vertebrate scale is ascended. In its development, the venous system of higher forms passes through certain stages common to the embryos of lower forms. The venous system consists of the sinus venosus (an accessory chamber present only in the hearts of lower vertebrates), cardinal veins, abdominal veins, coronary veins, pulmonary veins, and portal systems (in which veins break up into capillary networks before the blood coursing through them is returned to the heart). Note that all vertebrates have a hepatic portal system in which blood collected from the digestive tract and spleen passes through capillaries (sinusoids) in the liver before reaching the heart (Fig. 2).
Circulatory physiology
Circulatory physiology describes the structure and operation of the circulation in living animals, and it investigates how or why the circulatory system may have evolved. Comparing the circulatory system in different animal groups leads to an understanding of general principles and also to various applications of those principles, adapting animals to a wide range of habitats. The circulatory system in all vertebrates has multiple functions, but all functions are involved in regulating the internal environment of the animal (promoting homeostasis). See also: Homeostasis
In all vertebrates, the circulatory system (Fig. 5) consists of a central pump, the heart, which drives a liquid transport medium, the blood, continuously around a closed system of tubes, the vascular system. The arterial portion of this system is divided into larger elastic and smaller resistance vessels (arterioles), which distribute blood to specialized regions or organs (such as the muscles, gut, and lungs). Then, the transfer of nutrients, oxygen, or waste products takes place across the walls of a fine network of microscopic capillaries. Blood from the capillaries passes through the venules (small venous vessels) into the main veins and returns to the heart. The arterioles, venules, and capillaries make up the microcirculation, which is arguably the most important role of the vertebrate circulatory system from a functional point of view.