
The process whereby one or more gene alleles present in one chromosome may be exchanged with their alternative alleles on a homologous chromosome to produce a recombinant (crossover) chromosome which contains a combination of the alleles originally present on the two parental chromosomes. Genes which occur on the same chromosome are said to be linked, and together they are said to compose a linkage group. In eukaryotes, crossing-over may occur during both meiosis and mitosis, but the frequency of meiotic crossing-over is much higher. This article is concerned primarily with meiotic crossing-over. See also: Allele; Chromosome; Gene; Linkage (genetics)
Crossing-over is a reciprocal recombination event which involves breakage and exchange between two nonsister chromatids of the four homologous chromatids present at prophase I of meiosis; that is, crossing-over occurs after the replication of chromosomes which has occurred in premeiotic interphase. The result is that half of the meiotic products will be recombinants, and half will have the parental gene combinations (Fig. 1). Using maize chromosomes which carried both cytological and genetical markers, H. Creighton and B. McClintock showed in 1931 that genetic crossing-over between linked genes was accompanied by exchange of microscopically visible chromosome markers. See also: Recombination (genetics)
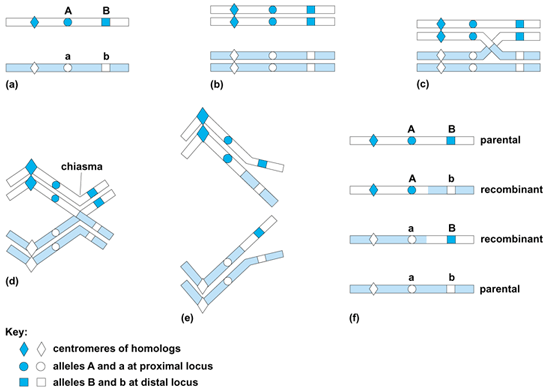
During meiosis, crossing-over occurs at the pachytene stage, when homologous chromosomes are completely paired. At diplotene, when homologs separate, the sites of crossing-over become visible as chiasmata, which hold the two homologs of a bivalent together until segregation at anaphase I. Each metaphase I bivalent will necessarily have at least one chiasma. In favorable material, such as grasshopper spermatocytes, it is possible to observe that each diplotene chiasma involves a crossover of two of the four chromatids at one site.
Where two or more crossovers occur in one bivalent, they usually do not cluster together but are widely separated; this is known as chiasma interference. The occurrence of one crossover event appears to preclude the occurrence of a second crossover in the immediate vicinity. In addition, the distribution of occurrence of chiasmata along a chromosome may be localized; the probability that a crossover will occur is higher in some chromosome segments and lower in other segments.
In general, the closer two genes are on a chromosome, that is, the more closely linked they are, the less likely it is that crossing-over will occur between them. Thus, the frequency of crossing-over between different genes on a chromosome can be used to produce an estimate of their order and distances apart; this is known as a linkage map. See also: Genetic mapping
Molecular mechanisms
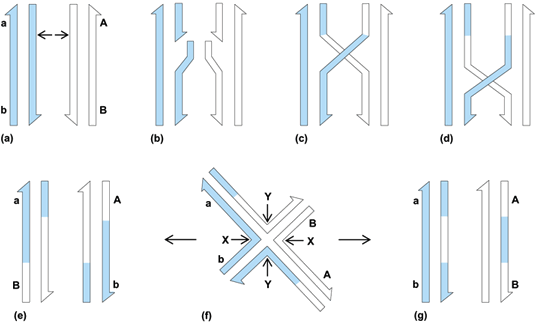
Since each chromatid is composed of a single deoxyribonucleic acid (DNA) duplex, the process of crossing-over involves the breakage and rejoining of DNA molecules. Although the precise molecular mechanisms have not been determined, it is generally agreed that the following events are necessary: (1) breaking (nicking) of one of the two strands of one or both nonsister DNA molecules; (2) heteroduplex (hybrid DNA) formation between single strands from the nonsister DNA molecules; (3) formation of a half chiasma, which is resolved by more single-strand breakages to result in either a reciprocal crossover, a noncrossover, or a nonreciprocal crossover (conversion event).
Two molecular models of recombination which have gained credence are those of R. Holliday and of M. Meselson and C. Radding. Holliday's model postulates nicks in both chromatids at the initiation of crossing-over (Fig. 2). Meselson and Radding postulate single-strand cut in only one DNA strand. Repair synthesis displaces this strand, which pairs with its complement on the other chromatid, thereby displacing and breaking the other strand of that DNA molecule. Following pairing and ligation of the two remaining broken ends, a half chiasma is formed. Other models have been postulated in which recombination is initiated by a double-stranded break in one chromatid. In all the above models, gene conversion can occur in the middle region of the molecules (with or without outside marker crossing-over) by mismatch repair of heteroduplex DNA.
Ultrastructural cytology
Pachytene, the meiotic stage at which crossing-over is considered to occur, corresponds with the period of close pairing or synapsis of homologous chromosomes. Electron microscopy has revealed that proteinaceous structures, the synaptonemal complexes (Fig. 3), are involved in the synapsis of chromosomes. A synaptonemal complex forms during zygotene by pairing of axial elements from two homologous chromosomes. It is present along the whole length of each pachytene bivalent and disappears at diplotene. Evidence from inhibitor studies and mutant stocks shows that the synaptonemal complex is necessary for meiotic crossing-over to occur. However, in cases such as desynaptic mutants, some hybrids, and the female silkworm, complete pachytene synaptonemal complexes have been observed, but no crossing-over occurs, showing that the synaptonemal complex alone is not sufficient to cause crossing-over.
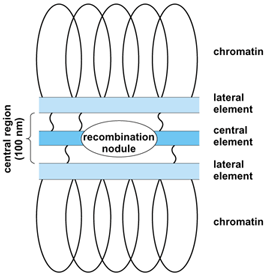
In Drosophila melanogaster oocytes, the occurrence at pachytene of dense spherical bodies bridging the central region of the synaptonemal complex has been described. These bodies coincided in number and position with expected crossover events, and therefore were named recombination nodules. A variety of oval and bar-shaped recombination nodules (Fig. 3) have also been found in organisms as diverse as fungi, humans, rat, silkworm, and maize. In many cases their number correlates with crossover frequency. It has been suggested that recombination nodules are prerequisites for crossing-over. If this is so, the recombination nodule may represent a complex of enzymes involved in the early events of recombination (nicking, strand separation, repair synthesis).
DNA repair synthesis has been observed during pachytene in lily microsporocytes, and has been shown to be reduced in an achiasmatic mutant. Prophase I of lilies is characterized by the presence of several proteins which could have a role in crossing-over, for example, DNA binding protein, endonucleases, ligases, and kinase. Inhibition of protein synthesis at zygotene-pachytene results in failure of crossing-over. Thus both DNA synthesis and protein synthesis appear necessary for meiotic crossing-over in lily. See also: DNA repair; Meiosis; Mitosis
Sex chromosomes
The differentiated X and Y sex chromosomes in human males and many animals (Z and W chromosomes in female birds) have small regions near one tip which undergo pairing and crossing-over at meiotic prophase I. Electron microscopy of the pachytene XY reveals the formation of a short synaptonemal complex segment with a recombination nodule in the majority of cases; the presence of a chiasma between the X and Y at metaphase I indicates the occurrence of crossing-over. An obligatory crossover in the XY bivalent is necessary to ensure regular segregation of X and Y to opposite poles at anaphase I. The pairing region contains a few gene loci on both X and Y chromosomes which exhibit an autosomallike inheritance pattern. Recombination between genes and DNA sequences in this pseudoautosomal region confirms the occurrence of obligatory crossing-over. The rare occurrence of XX males in some cases is accounted for by abnormal recombination events outside the pseudoautosomal region which have transferred the male sex-determining gene from the Y to the X chromosome. See also: Sex determination; Sex-linked inheritance

