
Key Concepts
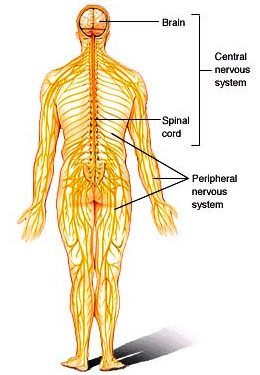
A coordinating and integrating system that functions in the adaptation of a vertebrate organism to its environment. The assemblages of cells that are specialized by their shape and function to act as the major coordinating organ of the body comprise the nervous system. Nervous tissue underlies the ability to sense the environment, to move and react to stimuli, and to generate and control all behavior of the organism. In general, an environmental stimulus causes a response in an organism when specialized structures (receptors) are excited. Excitations are conducted by nerves to effectors that act to adapt the organism to the changed conditions of the environment. With regard to the vertebrates, the nervous system (Fig. 1) consists of the brain, brainstem, spinal cord, cranial and peripheral nerves, and ganglia. The brain and spinal cord together comprise the central nervous system, whereas the nerves and ganglia that leave the brain and spinal cord constitute the peripheral nervous system. See also: Brain; Central nervous system; Cranial nerve; Ganglion; Nerve; Sensation; Sense organ; Spinal cord
Comparative morphology
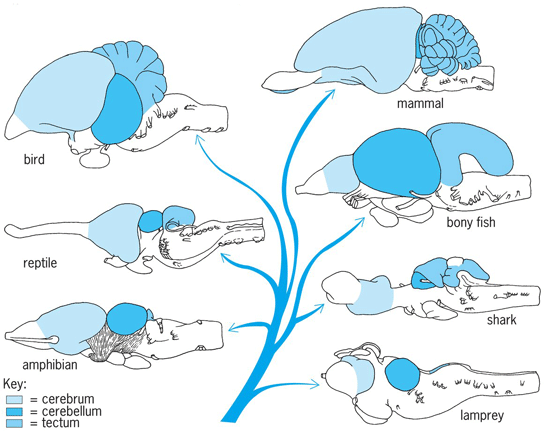
The brain of all vertebrates, including humans, consists of three basic divisions: prosencephalon, mesencephalon, and rhombencephalon (Fig. 2). The indication is that these divisions of the vertebrate brain have evolved along several functional lines and perform very different functions. See also: Animal evolution; Vertebrate brain
The nervous system of ancestral vertebrates apparently consisted of a single hollow neural tube (dorsally placed and running the length of the animal), which was probably structured in a manner similar to the spinal cord in living vertebrates (Fig. 3). It consisted of a central gray region that contained most of the cell bodies of the neurons (nerve cells) and was surrounded by a superficial white region containing the axons and dendrites of the neurons running to and from the tube. Each region of the tube was primarily concerned with the functions of that immediate region of the body to which its nerves projected. Neural functions were mediated at a segmental level rather than specialized in one specific region. Each region of the neural tube also contained neural cells that interconnected adjacent regions. This primitive condition still persists in a few living forms. See also: Neuron
As vertebrates evolved, structures in the head region became specialized for sensing the outside world and capturing food. This specialization was reflected in the neural tube, and its anterior region enlarged to permit analysis of the environment and integration of behavior. This enlargement of the neural tube is called a brain, and its divisions represent regions of different specialization. Each of the three primitive brain divisions was concerned with analysis and integration of a single type of sensory information. The rhombencephalon analyzed changes in the flow and pressure of water on an animal; the mesencephalon analyzed changes in the pattern and intensity of light; and the prosencephalon analyzed changes in the chemical composition of water. All vertebrates use these three types of information as clues in regulating their behavior. The individual divisions or patterns of the brain do not function separately to bring about a final response; rather, each pattern acts on a common set of connections in the spinal cord.
Spinal patterns
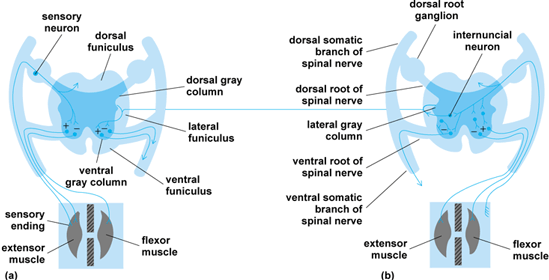
Spinal patterns are the final common patterns used by all higher brain pathways to influence all organs of the body. These reflexes are divided into two basic patterns: the monosynaptic arc and the multisynaptic arc (Fig. 3). The monosynaptic arc, or myotatic reflex, maintains tonus and posture in vertebrates and consists of two neurons—a sensory neuron and a motor neuron. The multisynaptic arc, or flexor reflex, is the pattern by which an animal withdraws a part of its body from a noxious stimulus. Both sensory neurons and internuncial neurons send information to brain centers (Fig. 3). Coordinated limb movement is based on a connective pattern of neurons at the spinal level. See also: Postural equilibrium
The structure of the spinal cord and its connections are basically similar among all vertebrates, and myotatic and flexor reflexes occur in all vertebrates. The major evolutionary changes in the spinal cord have been the increased segregation of cells and fibers of a common function from cells and fibers of other functions, and the increase in the length of fibers that connect brain centers with spinal centers.
Medullar patterns
The rhombencephalon of the brain is subdivided into a roof, or cerebellum, and a floor, or medulla oblongata. The medulla is similar to the spinal cord and is divided into a dorsal sensory region and a ventral motor region. It is an integrating and relay area between higher brain centers and the spinal cord. In addition to these nuclei and their connections, the medulla consists of both ascending and descending pathways to and from higher brain centers. The same basic connections occur throughout vertebrates.
Cerebellar patterns
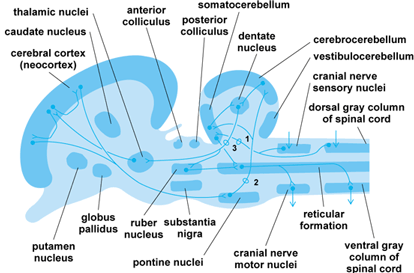
In all vertebrates, the cerebellum is divided into two major divisions: the two lateral flocculonodular lobes, and a central corpus cerebelli. The flocculonodular lobes are functionally referred to as the vestibulocerebellum and regulate vestibular reflexes underlying posture. The corpus cerebelli is subdivided into two lateral zones (the cerebrocerebellum) and a central zone (the somatocerebellum). In mammals, the lateral zones regulate corrective reflexes of posture and time muscular contractions of voluntary actions. The medial zone in mammals regulates reflex tonus of postural muscles by acting on the myotatic reflex. Note that, in mammals, the cerebellum does not initiate movement; it only times the length of muscle contractions and orders the sequence in which muscles should contract to bring about a movement. The command to initiate a movement is received from the cerebral cortex via the middle cerebellar peduncle (Fig. 4). Similarly, the cerebral cortex receives information regarding limb position and the state of muscular contraction to ensure that its commands can be carried out by the cerebellum.
Tectal patterns
The mesencephalon is divided into a roof, or optic tectum, and a floor, or tegmentum. The tegmentum contains the nuclei of the oculomotor and trochlear cranial nerves and a rostral continuation of the sensory nucleus of the trigeminal cranial nerve. Two motor nuclei dominate the tegmentum: the rubber nucleus and the substantia nigra. These nuclei are elements in the telencephalic and cerebellar motor systems.
Diencephalic patterns
In the evolution of vertebrates, the prosencephalon develops as two major divisions: the diencephalon and the telencephalon. The diencephalon retains the tubular form and serves as a relay and integrating center for information passing to and from the telencephalon and lower centers. The telencephalon is divided into a pair of cerebral hemispheres and an unpaired telencephalon medium.
There are three divisions of the diencephalon in all vertebrates: an epithalamus, which forms the roof of the neural tube; a thalamus, which forms the walls of the neural tube; and a hypothalamus, which forms the floor of the neural tube. The epithalamus and hypothalamus are primarily concerned with autonomic functions, including homeostasis. The thalamus is subdivided into dorsal and ventral regions. The dorsal region relays and integrates sensory information, whereas the ventral thalamus relays and integrates motor information. See also: Homeostasis; Instinctive behavior
Telencephalic patterns
The telencephalon is the most complex brain division in vertebrates. It is divided into a roof, or pallium, and a floor, or basal region. The pallium is divided into three primary divisions: a medial PI or hippocampal division; a dorsal PII or general pallial division; and a lateral PII division, often called the pyriform pallium. The basal region is divided into three areas: the first is a medial BIII area called the septum, whereas a ventral BII area and a lateral BI area form a region often called the corpus striatum.
The most striking change in the telencephalon of land vertebrates involves the PIIIa component, often called the dorsal ventricular ridge. In reptiles and birds, this region has proliferated into the ventricle of the telencephalon to produce a large cellular mass. In mammals, it has proliferated with the PIIb component of the dorsal pallium to produce the mammalian neocortex. In all land vertebrates except amphibians, the PIIb and the PIIIa components, along with the corpus striatum (BI and BII), are the highest centers for the analysis of sensory information and motor coordination. The PI, PIIa, PIIIb, BIII, and posterior parts of BI and BII form part of the limbic system, which is concerned with behavioral regulation.
Comparative histology
The basic microscopic anatomy of nervous tissue is essentially similar in all vertebrates, although variations do exist. Nervous tissue has the quality of irritability. It is also characterized by the quality of conductivity, which conveys the resulting excitation to other structures of the nervous system. The functional roles of nervous tissue include the capabilities to sense, through specialized receptors, environmental energies both internal and external to the organism; to conduct the resulting nerve impulses as coded input to centers in the nervous system; to process this input within these centers; to generate sensations and psychological expressions; and to produce such active responses as the contraction of muscles or the secretions of glands. In effect, nervous tissue reacts to environmental stimuli and regulates many bodily processes so as to maintain functional integrity of an organism. It is within the morphological, physiological, and chemical matrices of nervous tissue that the substrates for memory, behavior, and personality reside. See also: Learning; Memory; Personality theory
Neuron
Each neuron (Fig. 5), or nerve cell, possesses three fundamental properties, involving specialized capacity to react to stimuli, to transmit the resulting excitation rapidly to other portions of the cell, and to influence other neurons, muscles, or glandular cells. Each neuron consists of a cell body (soma), one to several cytoplasmic processes called dendrites, and one process called an axon. Cell bodies vary from about 7 to more than 70 micrometers in diameter; each contains a nucleus and several cytoplasmic structures, including Nissl (chromophil) granules, mitochondria, and neurofibrils. The cell body is continuously synthesizing new cytoplasm, especially protein, which flows down the cell processes. The dendrites range from a fraction of a millimeter to a few millimeters in length. An axon may range from about a millimeter up to many feet in length. The site where two neurons come into contact with each other and where influences of one neuron are transmitted to the other neuron is called a synapse. See also: Cytoplasm; Protein
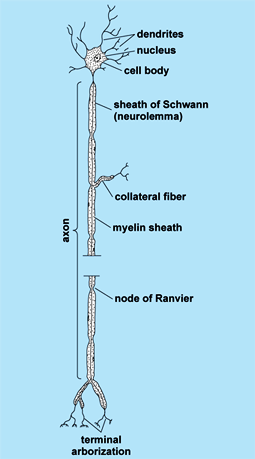
The cell membrane of the axon at the synapse is called the presynaptic membrane and that of the cell body or dendrite is called the postsynaptic membrane. The axon contains vesicles in the vicinity of the presynaptic membrane called presynaptic vesicles, which contain precursors of the neurotransmitter chemicals (for example, acetylcholine). Neurotransmitters are secreted across the presynaptic membrane into the synaptic cleft, where they may excite (excitatory synapse) or inhibit (inhibitory synapse) the postsynaptic membrane. Although a nerve fiber may transmit an impulse in either direction (toward or away from the cell body), conduction in a sequence of neurons is unidirectional. The impulse moves toward the cell body through dendrites and away from the cell body through the axon. This direction is established because the presynaptic neuron can stimulate the postsynaptic neuron, but the postsynaptic neuron cannot stimulate the presynaptic neuron. In this context, the synapse acts as a one-way valve and thereby establishes the functional polarity of the neuron. The site of contact of a nerve with a muscle, the motor end plate, is actually a synapse between a nerve and a muscle cell. See also: Acetylcholine; Cell membrane; Muscle; Synaptic transmission
Connective tissue cells
Nerve fibers of a peripheral nerve are bound together into small bundles by connective tissue cells, fibroblasts, and their fibrous products. The entire nerve is in turn surrounded by more connective tissues, within which are plexuses of blood vessels. Although the connective tissues are sparse within the central nervous system, the blood plexuses of the brain and spinal cord are most extensive. There are three layers of connective tissue membranes—the meninges—covering the brain and spinal cord: the innermost layer, the pia mater; the middle layer, the arachnoid; and the outermost layer, the dura mater. Between the pia mater and the arachnoid is the subarachnoid space; this space and the ventricular cavities within the brain are filled with an extracellular fluid called the cerebrospinal fluid. See also: Meninges
Comparative embryology
The anlage (primordium) of the nervous system is formed in the outer germ layer, the ectoderm, although some later contributions are also obtained from the middle germ layer, the mesoderm. In most vertebrates, a neural plate is formed, which later folds into a neural groove and then closes to form a neural tube. The formation of neural tissue within the ectoderm is due to inductive influences from underlying chordomesodermal structures. See also: Developmental biology; Embryonic induction; Germ layers; Morphogenesis; Neurulation
Neuromeres
When the neural tube is developing, a segmentation of the central nervous system occurs by the formation of transverse bulges, called neuromeres (Fig. 6). They are most distinctly seen in the hindbrain region, but can usually be identified in suitable embryonic stages in all parts of the central nervous system. Neuromeres are most easily seen in vertebrate brains having a thin wall, for instance, those of sharks, birds, reptiles, and mammals.
At the time of neuromeric segmentation, the brain is subdivided into the so-called brain vesicles by local widenings of its lumen. In the rostral end, more or less well-developed hemispheres are formed; in the middle of the brain anlage, the mesencephalic bulge develops; and behind the latter, the walls of the tube thicken into cerebellar folds. In this way, the brain anlage is divided into five sections (the telencephalon, diencephalon, mesencephalon, metencephalon, and myelencephalon), and its cavity is divided into the rudiments of the adult ventricles. The brain vesicles make the segmental characters of the neuromeric bulges less conspicuous.
Brain
In spite of the extraordinary variation in adult morphology of the vertebrate brain in different species, the early phases of development are essentially similar. The brain vesicle stages of a reptile, bird, and mammal are much alike; however, owing to varying growth rates of different parts and to specialization processes, the different patterns of the adult brains are formed.
Spinal cord
The spinal cord remains as a comparatively slightly differentiated tube. The primary lumen is secondarily reduced by the fusion of the side walls into a narrow central canal.
Cranial nerves
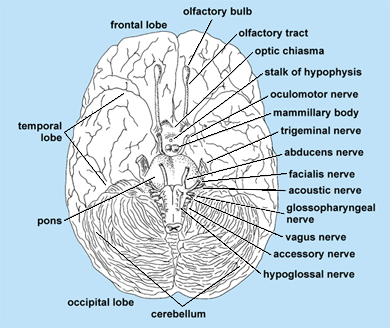
The cranial or cerebral nerves are the peripheral nerves of the head that are related to the brain. The number of nerves and the degree of development of the nerves vary in different species. The functional quality of the different nerves also varies. Twelve pairs of cranial nerves are distinguished in human anatomy (Fig. 7), and they are numbered rostrally to caudally as follows: (I) olfactory nerve, fila olfactoria; (II) optic nerve, fasciculus opticus; (III) oculomotor nerve; (IV) trochlear nerve; (V) trigeminal nerve (in most vertebrates, this is divided into three branches: ophthalmic, maxillary, and mandibular); (VI) abducens nerve; (VII) facial nerve; (VIII) statoacoustic nerve; (IX) glossopharyngeal nerve; (X) vagus nerve; (XI) accessory nerve; and (XII) hypoglossal nerve. The varied morphological significance of the cranial nerves is evident from their embryology.
Spinal nerves
The spinal ganglia are formed from the neural crest, which grows out like a continuous sheet from the dorsal margin of the neural tube and is secondarily split up into cell groups (the ganglia) by a segmentating influence from the somites. Fibers grow out from the ganglionic cells and form the sensory fibers of the spinal nerves. Motor nerve fibers emerge from cells situated in the ventral horns of the spinal cord. The ventral motor fibers and the dorsal sensory fibers fuse to form a common stem, which is again laterally divided into branches, innervating the corresponding segment of the body. See also: Neural crest
Autonomic nervous system
The autonomic nervous system functions primarily at a subconscious level. It is traditionally partitioned into the sympathetic system and the parasympathetic system. The ganglia of the sympathetic nervous system develop ventrolateral to the spinal cord as neural crest derivatives. At first, a continual column of sympathetic nerve cells is formed; it later subdivides into segmental ganglia. The parasympathetic system is made up of preganglionic fibers emanating as general visceromotor fibers from the brain and from sacral segments of the spinal cord. Cells migrate to form the peripheral ganglia along them. See also: Autonomic nervous system; Parasympathetic nervous system; Sympathetic nervous system

