
Key Concepts
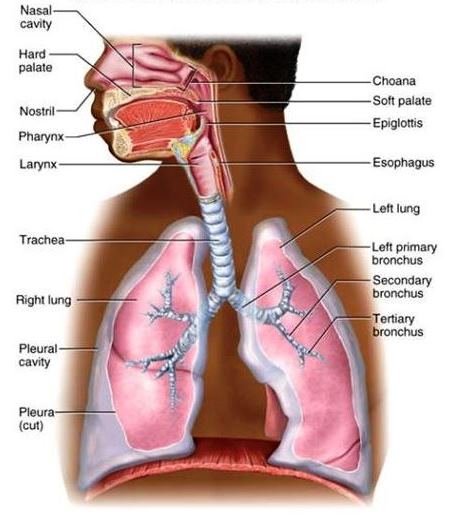
The system of organs involved in the acquisition of oxygen and the elimination of carbon dioxide by an organism. The respiratory system consists of a set of specialized organs, including anatomical structures and passages, that are responsible for the proper intake and exchange of oxygen and carbon dioxide between an organism's body and the environment. Structurally, respiratory organs usually present a vascular surface that is sufficiently extensive to provide an adequate area of absorption for gaseous exchange. This surface is moist and thin enough to allow for the passage of gases. Across the diversity of life, various respiratory structures have developed, including the lungs and airways of air-breathing vertebrates (Fig. 1), the gills of fishes and many invertebrates, and the specialized air ducts (tracheae) of insects. With regard to vertebrates, the lungs and gills are the two most important structures involved in the phase known as external respiration, or gaseous exchanges, between the blood and environment. Internal respiration refers to the gaseous exchanges that occur between the blood and cells. Certain other structures in some species of vertebrates serve as respiratory organs—for example, the integument or skin of fishes and amphibians. In particular, the moist, highly vascular skin of anuran amphibians is important in respiration. In addition, certain species of fishes have a vascular rectum that is utilized as a respiratory structure (with water being taken in and ejected regularly by the animal), whereas saclike cloacal structures occur in some aquatic species of turtles. These latter structures are vascular and are intermittently filled with, and emptied of, water; it is thought that they may function in respiration. It also should be mentioned that the yolk sac and allantois are important respiratory organs during the embryonic life of certain vertebrates. See also: Allantois; Blood; Carbon dioxide; Lung; Oxygen; Respiration; Skin; Vertebrata; Yolk sac
Gills
Gills are involved in many functions, including the exchange of oxygen, carbon dioxide, water, acids, inorganic ions, ammonia, and urea between the body and the surrounding environment (water). Much of the transfer of these compounds is passive, but some ions are moved against an electrochemical gradient requiring the expenditure of energy. As a result, the gills are a metabolically active tissue; they consume up to 10–25% of the oxygen uptake of a resting fish. See also: Energy metabolism; Metabolism
Functionally, the gills can be considered as a large, thin, epithelial surface layer separating the inside (blood) of the animal from the water. They are specialized for the exchange of material in water (in contrast to lungs, which have evolved for gas exchange in air). To ensure adequate oxygen transfer from water into the animal, most aquatic animals maintain a continual, unidirectional flow of water over the gills, with only very small diffusion distances. The gills are essentially a fine sieve placed in the path of the water flow so that a thin layer of water passes over each region of the gill epithelium.
Fish
The exchange units of the gills of fishes are the lamellae, which are arranged in rows on filaments attached to gill arches (Fig. 2). Blood flows through the lamellae in the opposite direction to water flow, creating a countercurrent exchange system between blood and water across the gill epithelium. Oxygen is delivered to the gill surface by water flow, diffuses across the gill epithelium, and is carried away in the blood bound to hemoglobin. Oxygenated blood (because of the presence of hemoglobin) contains much more oxygen per unit volume than water. See also: Countercurrent exchange (biology); Hemoglobin
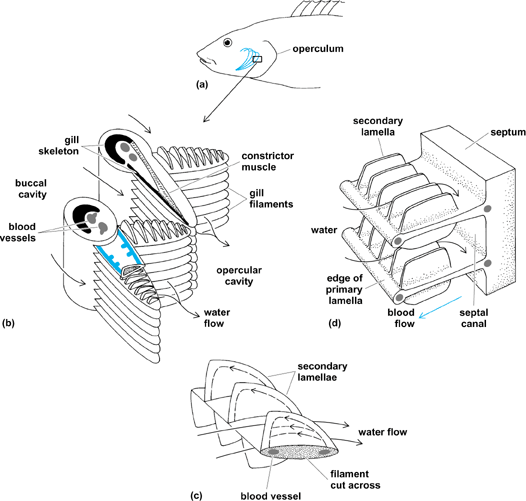
Water flow over the gills of fishes is maintained by the action of muscular pumps and unidirectional flap valves situated at the mouth and the opercular cleft in bony fishes, and at the mouth and gill slits of elasmobranches. The valves are skin folds that permit flow in only one direction, into the mouth and out via the gill slit. The mouth (buccal) cavity expands, drawing water into the mouth; at the same time, the postgill or opercular cavity expands with the outlet valve closed, drawing water over the gills; the opercular and buccal cavities then contract, forcing water out via the gill slit. The buccal force and opercular suction pumps operate at the same frequency, but slightly out of phase; thus, although water velocity over the gills varies with each breathing cycle, there is nearly always some water flow over the gills, with little or no backflow. The relative importance of the buccal versus the opercular pump differs among species.
Low oxygen levels often occur in rivers and lakes and even in the sea. Under these conditions, many fish supplement their oxygen supply by breathing air. The air-breathing organ is usually the swim bladder or a buccal or gut cavity, and not the gills, which are retained for gas transfer in water. Some fish are obligate air breathers and die if denied access to air. In this case, the gills are retained for excreting carbon dioxide into water and maintaining ionic balance. The air-breathing organ is for oxygen uptake. This is associated with changes in the circulatory system. See also: Circulation; Swim bladder
Amphibians
Almost all amphibians have gills at some time during development and may retain them as adults. In some amphibians, the gill structures may be external, that is, the gills are not enclosed in a buccal cavity (Fig. 3). These gills are not ventilated, but are waved around in the water to ensure adequate oxygen delivery. Other amphibians have neither gills nor lungs, but obtain oxygen from the water simply by diffusion across the general body surface. See also: Diffusion

Lungs
The lung is an organ adapted to the respiratory exchange of gases between air and the blood of vertebrates. The shape and volume of the lung (Fig. 4), because of its pliability, conforms almost completely to that of its cavity. The lungs are conical; each has an apex and a base, two surfaces, two borders, and a hilum. The apex extends into the superior limit of the thoracic cavity. The base is the diaphragmatic surface. The costal surface may show bulgings into the intercostal spaces. The medial surface has a part lying in the space beside the vertebral column and a part imprinted by the form of structures bulging outward beneath the mediastinal pleura. The hilum and pulmonary ligament descending from it are notable. The cardiac impression is deeper on the left lung because of the position of the heart. Essential to lung function, histologically, are the respiratory passages, pleura, blood vessels, lymphatics, and respiratory surfaces proper. See also: Lung
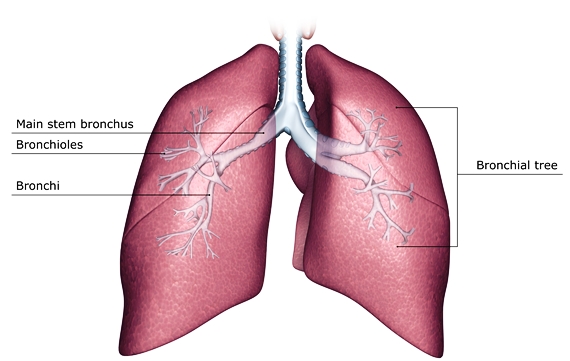
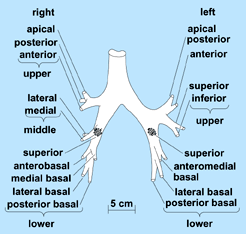
For convenience, the lung may be divided into anatomical areas. The bronchial tree branches mainly by dichotomy (Fig. 5). The ultimate generations, that is, the respiratory bronchioles, alveolar ducts, and alveoli, constitute all of the respiratory portion of the lung. This respiratory portion consists of 10 segments in the right lung and 8 in the left, with each being supplied by a tertiary branch of the bronchial tree (Fig. 5). Two or more of the segments make up a lung lobe. Furthermore, the primary bronchi (from the trachea) divide as secondary bronchi, with three on the right side and two on the left, corresponding to the lobes of the lung. Because the upper left lobe results from the fusion of two lobes, the prospective left upper and middle lobar bronchi become partially fused. As a result, the four segmental bronchi of the upper lobe are not tertiary, but bronchi of the fourth division of the tree. However, there is great similarity in the structure of the right and left bronchi.
The blood supply to the lung is provided by the pulmonary and the bronchial arteries. The nerves that supply the lung are branches of the vagus and of the thoracic sympathetic ganglia 2, 3, and 4. Efferent vagal fibers have bronchoconstrictor and secretory functions, whereas the afferents are part of the arc for the breathing reflex. Efferent sympathetic fibers are bronchodilators; hence, adrenaline is used for relief of bronchial spasm resulting from asthma. See also: Asthma; Nervous system (vertebrate)
In addition, the evolutionary history of the lung includes the development of certain vital accessories to respiration, specifically, the diaphragm and thoracic wall. All lungs correspond in originating as a pocket from the pharynx near the level of the sixth aortic arches, which furnish the blood supply; in innervation by the vagus nerve; and in their embryonic position in the abdomen. This position is retained in the adult forms of fish, amphibians, and reptiles and is partially retained in birds; however, in mammals, the lung is segregated from the abdomen in fetal life into a new-formed thoracic cavity (Fig. 6). See also: Pharynx
Vertebrate respiratory regulation
The ventilation and circulation through the lungs of vertebrates are controlled and adjusted to maintain an efficient exchange of gases in relationship to the needs of the animal and to the suitability of the environment. Sensory units (tension receptors) for pressure or distension occur in lung and breathing elements and in related circulatory organs. These signal the medulla oblongata of the brain, from which the breathing rhythm itself originates, as well as other parts of the brain, and breathing is adjusted to physiological changes and physical conditions. Other receptors, especially in certain major blood vessels near the heart and in the medulla itself, signal the respiratory areas of the brain, providing information about blood chemical conditions (in particular, oxygen, carbon dioxide, and acidity information). Signals from other parts of the body (for example, the olfactory and tracheal membranes and the joints) and from other parts of the brain (including the heat-regulating, olfactory, and phonation areas) also reach respiratory areas. Here, the correlating and integrating role of the brain determines how breathing will proceed: whether to speed up, stop, cough, or do whatever is appropriate. Oxygen tension in blood is a basic regulatory element for the maintenance of the ventilation rate in lower vertebrates, but it becomes secondary to carbon dioxide in birds and mammals. Because carbon dioxide is a major factor in blood acidity, ventilation also comes to play a major role in the critical regulation of hydrogen ion concentration (blood acidity) in higher animals. See also: Brain; pH regulation (biology)
Adaptations in terrestrial animals
The adaptations for diving in birds and mammals have been found in counterpart among some nonaquatic animals, such as rabbits, which may become immobile and suppress breathing under certain conditions, for example, in evading predators. For active locomotion in air and for highest brain activity, however, oxygen is the most urgently needed of all substances required from the environment; therefore, essentially continuous ventilation is a necessity for strictly terrestrial animals (including, for example, chickens, cats, and humans). Such animals survive without actual ventilation only before the time of birth, or hatching, although nonrespiratory periodic breathing movements do occur in the egg or uterus as appropriate structures develop. Early respiration utilizes special diffusion respiratory structures, including the allantois and placenta. In birds, an actual pulmonary ventilation, utilizing the air chamber of the egg, begins shortly before hatching. In mammals, it can begin only when the fetus gains direct access to air at birth. See also: Adaptation (biology); Diving animals; Placentation

