
Key Concepts
The microscopic functional and structural unit of all living organisms. Cells can be separated into prokaryotic and eukaryotic categories. Eukaryotic cells contain a nucleus. They comprise protists (single-celled organisms), fungi, plants, and animals, and they are generally 5–100 micrometers (μm) in linear dimension. Prokaryotic cells contain no nucleus, are relatively small (1–10 μm in diameter), and have a simple internal structure (Fig. 1). They include two classes of bacteria: eubacteria (including photosynthetic organisms, or cyanobacteria), which are common bacteria that inhabit soil, water, and larger organisms; and archaebacteria, which grow under unusual conditions. The archaebacteria include methanogens, which live in anaerobic conditions and reduce carbon dioxide (CO2) to methane (swamp gas); halophiles, which thrive under extremely high salt conditions; and sulfur bacteria, which grow in hot (80°C; 176°F) sulfur springs, where the pH is extremely acidic. See also: Archaea; Bacteria; Cell biology; Eukaryota; Prokaryote
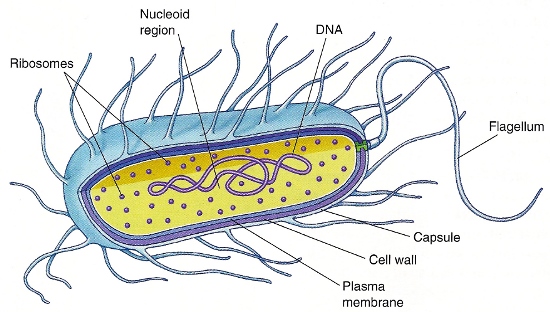
Prokaryotic (bacterial) cells
All eubacteria have an inner (plasma) membrane that serves as a semipermeable barrier (Fig. 1), which allows small nonpolar and polar molecules (for example, oxygen, carbon dioxide, and glycerol) to diffuse across (down their concentration gradients), but does not allow the diffusion of larger polar molecules (for example, sugars and amino acids) or inorganic ions (for example, Na+, K+, Cl−, and Ca2+). The plasma membrane, which is a lipid bilayer, utilizes transmembrane transporter and channel proteins to facilitate the movement of these molecules. See also: Cell membranes
Eubacteria can be further separated into two classes based on their ability to retain the dye crystal violet. Gram-positive cells retain the dye; their cell surface includes the inner plasma membrane and a cell wall composed of multiple layers of peptidoglycan. Gram-negative bacteria are surrounded by two membranes: the inner (plasma) membrane and an outer membrane that allows the passage of molecules of less than 1000-dalton (Da) molecular weight through porin protein channels. The peptidoglycan-rich cell wall and the periplasmic space are located between the inner and outer membranes.
Eubacteria contain a single circular double-stranded molecule of deoxyribonucleic acid (DNA), or a single chromosome. Because prokaryotic cells lack a nucleus, this genomic DNA resides in a central region of the cell (Fig. 1) called the nucleoid region. Bacterial replication requires accurate duplication of this chromosomal DNA, with both copies attaching by cross-wall formation to the plasma membrane, allowing the formation of two identical daughter cells by binary fission. The bacterial genome contains all the information necessary to maintain the structure and function of the cell. See also: Chromosome; Deoxyribonucleic acid (DNA)
Many bacteria are able to move from place to place, or are motile. Their motility is based on a helical flagellum composed of an interwoven protein called flagellin. The flagellum is attached to the cell surface through a basal body and propels the bacteria through an aqueous environment by rotating (this action is similar to that of a propeller on a motorboat). The movement is reversible, allowing the bacteria to move toward chemoattractants and away from chemorepellents. See also: Cilia and flagella
Eukaryotic cells
In a light microscopic view of a eukaryotic cell, a plasma membrane can be seen that defines the outer boundaries of the cell, surrounding the cell's protoplasm or contents. The protoplasm includes the nucleus, where the eukaryotic cell's DNA is compartmentalized away from the remaining contents of the cell (the cytoplasm); therefore, the protoplasm includes the nucleus and the cytoplasm. In addition, the cytoplasm contains other organelles (specialized subcellular structures that carry out a specific function), including the mitochondrion, endoplasmic reticulum, Golgi apparatus, lysosome, peroxisome, cytoskeleton, and plasma membrane (Fig. 2). The organelles occupy approximately half the total volume of the cytoplasm. The remaining compartment of cytoplasm (minus the organelles) is referred to as the cytosol or cytoplasmic ground substance. Eukaryotic cells also have a cytoskeleton that gives the cell its shape, its capacity to move, and its ability to transport organelles and vesicles from one part of the cell cytoplasm to another. The cytoskeleton is composed of microfilaments (7–8 nm in diameter), intermediate filaments (10 nm in diameter), microtubules (25 nm in diameter), and a spectrin-based membrane skeleton. Eukaryotic cells are generally larger than prokaryotic cells and thus require a cytoskeleton and membrane skeleton to maintain their shape. See also: Cell organization; Cytoplasm
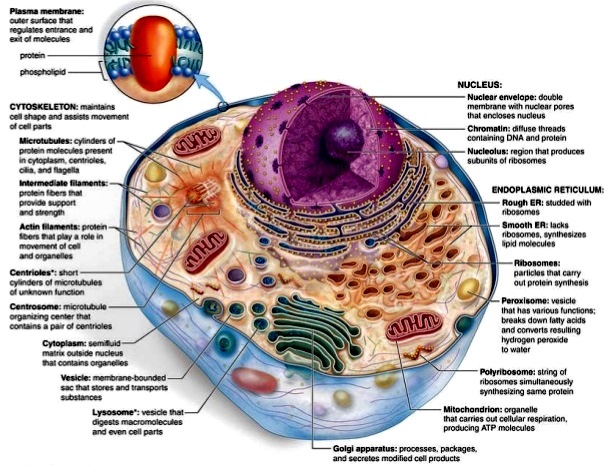
Eukaryotic cells contain a large amount of DNA (about 1000-fold more than bacterial cells), with only approximately 1% encoding protein. The remaining DNA is structural (involved in DNA packaging) or regulatory (helping to switch on and off genes). Therefore, whether a cell becomes a muscle fiber or a neuron depends on the expression of muscle-specific genes or neuron-specific genes (although both are present). See also: Gene; Protein
Plasma membrane
The plasma membrane serves as a selective permeability barrier between a cell's environment and its cytoplasm. The fundamental structure of plasma membranes (as well as organelle membranes) is the lipid bilayer, formed as a result of the tendency of amphipathic phospholipids to bury their hydrophobic fatty acid tails away from water. Human and animal cell plasma membranes contain a varied composition of phospholipids, cholesterol, and glycolipids. Although lipids determine the bilayer structure, membrane proteins are primarily responsible for membrane function. Both the lipids and proteins of the plasma membrane are distributed asymmetrically across the bilayer. This creates a membrane topography in which selected membrane domains can have differing protein and lipid composition and also function. The so-called lipid rafts constitute one of these domains, comprising a cholesterol-rich region of the lipid bilayer containing specific proteins (such as flotillin and stomatin). See also: Cholesterol; Lipid; Lipid rafts (membranes); Phospholipid
Cytoskeleton
The cytoskeleton is involved in establishing cell shape, polarity, and motility, as well as in directing the movement of organelles within the cell. The cytoskeleton includes microfilaments, microtubules, intermediate filaments, and the two-dimensional membrane skeleton that lines the cytoplasmic surface of cell membranes. See also: Cytoskeleton
Nucleus
One of the most prominent organelles within a eukaryotic cell is the nucleus. The nuclear compartment is separated from the rest of the cell by a specialized membrane complex built from two distinct lipid bilayers, referred to as the nuclear envelope. However, the interior of the nucleus maintains contact with the cell's cytoplasm via nuclear pores. These structures form holes or channels in the nuclear envelope that are responsible for the selective uptake of nuclear components and the release of components destined for the cell cytoplasm. The primary function of the nucleus is to house the genetic apparatus of the cell; this genetic machinery is composed of DNA (arranged in linear units called chromosomes), ribonucleic acid (RNA), and proteins. Nuclear proteins aid in the performance of nuclear functions and include polypeptides that have a direct role in the regulation of gene function and those that give structure to the genetic material. The complex of DNA and structural proteins is known as chromatin, which can be either highly compacted (heterochromatin) or dispersed (euchromatin) within the nucleus. A specialized region within the nucleus that is visible in interphase cells is the nucleolus, which functions in the production and assembly of ribosomal subunits. See also: Cell nucleus; Ribonucleic acid (RNA); Ribosomes
Endoplasmic reticulum
The endoplasmic reticulum is composed of membrane-enclosed flattened sacs or cisternae. The enclosed compartment is called the lumen. The endoplasmic reticulum (ER) is separated morphologically into a rough ER (RER) and a smooth ER (SER). The RER is studded with ribosomes, whereas the SER is not. The RER is the site of protein synthesis, whereas lipids are synthesized in the RER and the SER. See also: Endoplasmic reticulum
Golgi apparatus
The final modifications of proteins and glycolipids occur within a series of flattened membranous sacs comprising the Golgi apparatus. Vesicles that bud from the endoplasmic reticulum fuse with a specialized region of the cis Golgi compartment called the cis Golgi network. Modifications occur first in the cis compartment and then in the medial and trans Golgi network compartments. In each case, the membrane transfer occurs by the blebbing of coated vesicles from one compartment and their fusion with the next. In the trans Golgi network, proteins and lipids are sorted into transport vesicles destined for lysosomes, the plasma membrane, or secretion. See also: Golgi apparatus
Lysosome
The lysosome is a membrane-bound organelle with a luminal pH of 5.0 that is filled with acid hydrolases. Found in animal cells (and not plant cells), the lysosome is responsible for degrading materials brought into the cell by endocytosis or phagocytosis, or by autophagocytosis of spent cellular material. See also: Lysosome
Mitochondrion
The mitochondrion contains a double membrane: the outer membrane, which contains a channel-forming protein named porin; and an inner membrane, which contains multiple infolds called cristae. The porin channel allows molecules of less than 10,000-Da molecular weight free passage between the cytoplasm and the intermembrane space. The inner membrane, which contains the protein complexes responsible for electron transport and oxidative phosphorylation, is folded into numerous cristae that increase the surface area per volume of this membrane. The transfer of electrons from nicotinamide adenine dinucleotide (NADH) or flavin adenine dinucleotide (FADH2) down the electron-transfer chain to oxygen causes protons to be pumped out of the mitochondrial matrix into the intermembrane space. The resulting proton motive force drives the conversion of adenosine diphosphate (ADP) plus inorganic orthophosphate (Pi) to adenosine triphosphate (ATP) by the enzyme ATP synthetase. See also: Adenosine triphosphate (ATP); Mitochondria; Nicotinamide adenine dinucleotide (NAD)
Peroxisome
Within the peroxisome, hydrogen atoms are removed from organic substrates, and hydrogen peroxide is formed. The enzyme catalase can then utilize the hydrogen peroxide to oxidize various substrates, including alcohols, formaldehydes, and formic acid, in detoxifying reactions. See also: Peroxisome
Additional features of plant cells
Plant cells are distinguished from other eukaryotic cells by various features (Fig. 3). For example, plant cells have an extremely rigid cell wall outside their plasma membrane. This cell wall is composed of cellulose and other polymers and is distinct in composition from the cell walls found in fungi or bacterial cells. The plant cell wall expands during cell growth, and a new cell wall partition is created between the two daughter cells during cell division. Similar cell walls are not observed in animal cells. See also: Cell division; Cell walls (plant); Cellulose; Plant cell; Plant growth
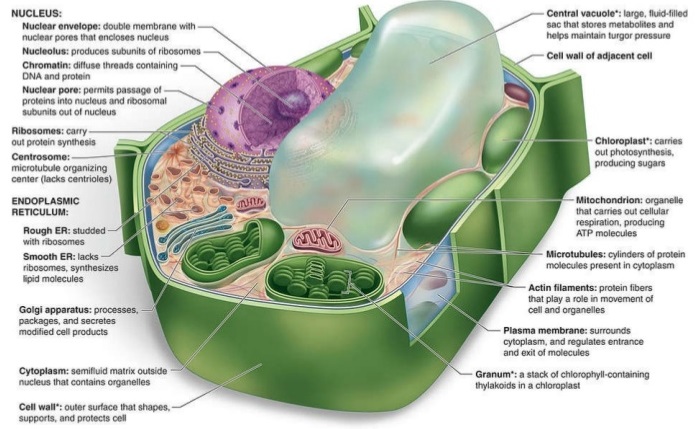
Most plant cells contain membrane-encapsulated vacuoles as major components of their cytoplasm. These vacuoles contain water, sucrose, ions, nitrogen-containing compounds formed by nitrogen fixation, and waste products. The amount of osmotic particles within the vacuoles is much higher than in the cytosol or extracellular fluid. The encapsulating membrane is permeable to water, but impermeable to the molecules within the vacuole. Therefore, water rushes into the vacuole, the vacuole expands, and a hydrostatic pressure or turgor is created within the cell. This turgor is resisted by the cellulose-containing cell wall; however, as the cell wall stretches, the plant grows. See also: Vacuole
Chloroplasts are the other major organelles in plant cells that are not found in other eukaryotic cells. Like mitochondria, they are constantly in motion within the cytoplasm. One of the pigments found in chloroplasts is chlorophyll, which is the molecule that absorbs light and gives the green coloration to the chloroplast. Chloroplasts, like mitochondria, have outer and inner membranes. Within the matrix of the chloroplast, there is an intricate internal membrane system. The internal membranes are made up of flattened interconnected vesicles, called thylakoid vesicles, which take on a disklike structure. The thylakoid vesicles are stacked to form structures called grana, which are separated by a space called the stroma. Within the stroma, carbon dioxide fixation occurs, in which carbon dioxide is converted to various intermediates during the production of sugars. Chlorophyll is found within the thylakoid vesicles; it absorbs light and, with the involvement of other pigments and enzymes, generates ATP during photosynthesis. See also: Cell plastids; Chlorophyll; Photosynthesis

