
Key Concepts
A set of structures and organs in which food substances are digested. The vertebrate digestive system consists of the digestive tract (alimentary canal) and ancillary organs that serve for the acquisition of food and the assimilation of nutrients required for energy, growth, maintenance, and reproduction. Food is ingested, reduced to particles, mixed with digestive fluids and enzymes, and propelled through the digestive tract. Enzymes produced by the host animal and microbes indigenous to the digestive tract destroy harmful agents and convert food into a limited number of nutrients, which are selectively absorbed. Some characteristics of the vertebrate digestive system are common to all classes of vertebrates and appear to have been conserved in their evolution. In other aspects, the digestive systems of vertebrates show numerous structural and functional adaptations to their diet, habitat, and other physiological characteristics (Fig. 1). Carnivores, which feed exclusively on other animals, and species that feed on plant concentrates (seeds, fruit, nectar, and pollen) tend to have the shortest and simplest digestive tract. The digestive tract tends to be more complex in omnivores, which feed on both plants and animals, and most complex in herbivores, which feed principally on the fibrous portions of plants. See also: Adaptation (biology); Animal evolution; Energy metabolism; Enzyme; Food; Nutrition; Physiological ecology (animal)
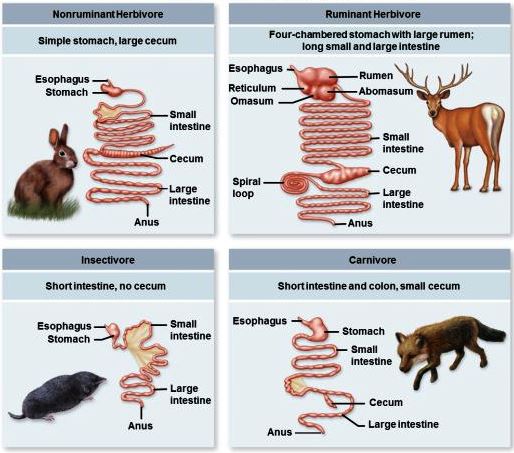
Anatomy
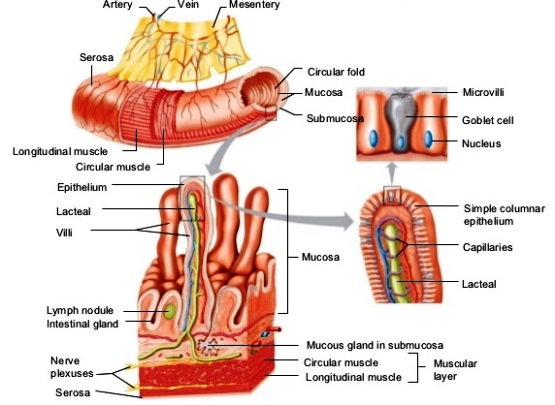
Because of wide species variations, the digestive system of vertebrates is best described in terms of the headgut, foregut, midgut, pancreas, biliary system, and hindgut. The various structures and organs associated with the human digestive system are shown in Fig. 2.
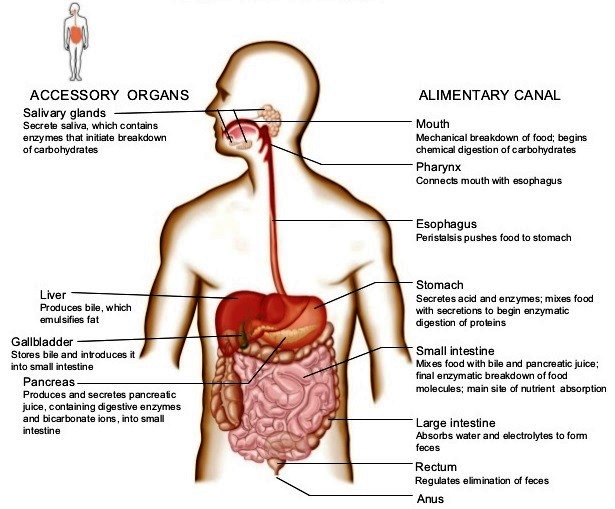
Headgut
The headgut consists of the mouthparts and pharynx, which serve for the procurement and the initial preparation and swallowing (deglutition) of food. Movable, articulated jaws are found in all vertebrates, except the cyclostomes (lampreys and hagfish), which are primitive, parasitic fish. Teeth, used for grasping, cutting, or tearing of food, are located in the jaws or other mouthparts of most fish, and adult amphibians are carnivores with a weak dentition that is used only for grasping and positioning prey. Most reptiles have teeth that are used for the procurement, cutting, tearing, or crushing of food, but turtles and birds use a beak for these purposes. A few mammals have no teeth; however, most mammalian species possess incisors and canines for grasping, cutting, and tearing, and premolars and molars with uneven occluding surfaces. Both the articulation and the musculature of the mammalian jaws allow the vertical movement of the lower jaw and either its lateral movement, as seen in most species, or a forward and backward movement, as seen in rodents and elephants. Thus, mammals have the unique ability to use their premolar and molar teeth for the grinding and crushing of food. A movable tongue aids in the procurement of food (for example, this is seen in frogs, toads, woodpeckers, and anteaters) and for the placement of food between the molars of mammals. In addition, oral glands secrete fluids that aid in the swallowing of food, as well as adhesive materials or even toxins in a number of species. See also: Mouth; Oral glands; Pharynx; Tongue; Tooth
Foregut
The foregut consists of an esophagus that is used for the swallowing of food and, in most species, a stomach (Fig. 3) that serves for its storage and initial stages of digestion. The esophagus of most vertebrates is lined with multilayers of cells that are impermeable to absorption. In most birds, it contains the crop, which is an outpocketing of its wall that provides for the temporary storage of food. A stomach is present in the overwhelming majority of vertebrates; the exceptions include the cyclostomes, some species of advanced fish, and the larval amphibians. In most vertebrates, it consists of a dilated segment of the gut that is separated from the esophagus and midgut by muscular sphincters or valves. This is often referred to as a simple stomach. However, in birds, these functions are carried out by the crop (storage), proventriculus (secretion), and gizzard (grinding or mastication). See also: Esophagus; Stomach
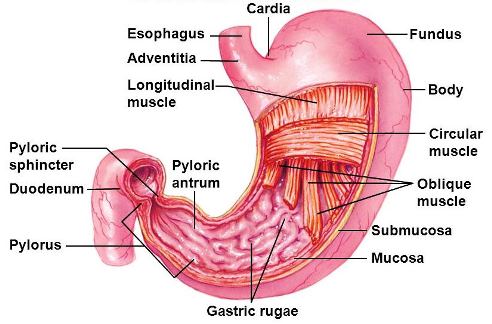
In most vertebrates, a major portion of the stomach is lined with a proper gastric mucosa (epithelium) [Fig. 4], which secretes mucus, hydrochloric acid (HCl), and pepsinogen. The distal (pyloric) part of the stomach secretes mucus and bicarbonate ions (HCO3−), and its muscular contractions help reduce the size of food particles and transfer partially digested food into the midgut. The stomach of reptiles and most mammals has an additional area of cardiac mucosa near its entrance, which also secretes mucus and bicarbonate ions. The stomach of some mammals differs from that of other vertebrates with respect to its epithelial lining, structure, or size.
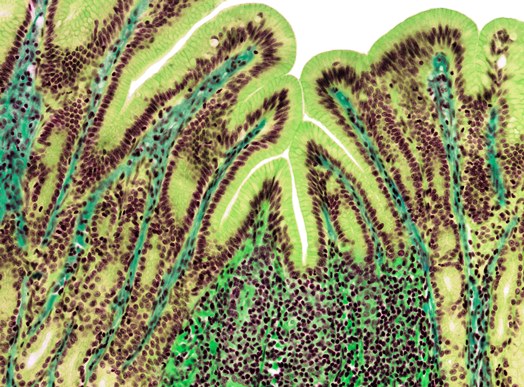
Midgut
The midgut or small intestine is the principal site for the digestion of food and the absorption of nutrients. It is lined with a single layer of cells that (1) secrete mucus and fluids, (2) contain enzymes that aid in the final stages of the digestion of carbohydrates and proteins, and (3) absorb nutrients from the lumen into the circulatory system. The surface area of the lumen can be increased by a variety of means. For example, in higher vertebrates, the lumen surface is increased by the presence of villi (Fig. 5), which are macroscopic projections of the epithelial and subepithelial tissues. A crypt at the base of each villus contains cells that secrete fluid into the gut and hormones that help regulate the functions of the digestive system. The crypts also contain cells that serve as precursors for the generation of absorptive and mucus-producing cells, which migrate up the villus surface and are eventually sloughed off into the lumen. See also: Intestine
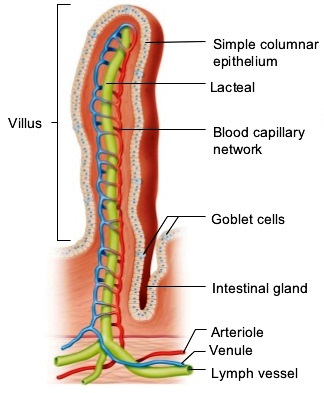
The lumen surface in all vertebrates is also expanded by a brush border of microvilli on the lumen-facing (apical) surface of the midgut absorptive cells (Fig. 6). The brush border membranes contain enzymes that aid in the final digestion of food and they assist in the selective absorption of nutrients. The lumenal surface area of the human small intestine is increased 10-fold by the presence of villi and an additional 20-fold by the presence of microvilli, resulting in a total surface area of 2,000,000 cm2 (310,000 in.2).

Pancreas and biliary system
Digestion in the midgut is aided by secretions of digestive enzymes and fluid by pancreatic tissue, as well as by the secretion of bile by the liver. Pancreatic tissue is represented by primitive ceca (blind sacs) along the midgut of cyclostomes and is distributed along the intestinal wall, and even into the liver, of some species in the more advanced classes of fish. However, the pancreas is a compact organ in sharks, skates, rays, many teleosts (ray-finned fishes), and all other vertebrates. The liver is another compact organ in all vertebrates. One of its many functions is the secretion of bile. In most vertebrates, the bile is stored in the gallbladder and released into the intestine as needed, but a gallbladder is absent in some species of fish and mammals. Bile salts serve to emulsify lipids and increase their surface area available for digestion by the water-soluble lipase. See also: Gallbladder; Lipid; Liver; Pancreas
Hindgut
The hindgut is the final site of digestion and absorption prior to defecation or evacuation of waste products. The hindgut of fish, amphibian larvae, and a few mammals is short and difficult to distinguish from the midgut. However, the hindgut of adult amphibians and reptiles, birds, and most mammals is a distinct segment, which is separated from the midgut by a muscular sphincter or valve. It also tends to be larger in diameter. Thus, the midgut and the hindgut of these animals are often referred to as the small intestine and the large intestine, respectively.
The hindgut of some reptiles and many mammals includes a blind sac (cecum) near its junction with the midgut. A pair of ceca are present in the hindgut of many birds and a few mammalian species. The remainder of the hindgut consists of the colon and a short, straight, terminal segment, which is called the rectum in mammals. The digestive and urinary tracts exit separately from the body of most species of fish and mammals. However, in adult amphibians and the reptiles, birds, and some mammals, this segment terminates in a chamber called the cloaca, which also serves as an exit for the urinary and reproductive systems. The hindgut or, if present, the cloaca terminates in the anus. See also: Colon; Urinary system
The hindgut is similarly lined with a single layer of absorptive and mucus-secreting cells that originate in crypts. However, it lacks villi, and (with the exception of the cecum of birds) its absorptive cells lack digestive enzymes and the ability to absorb most nutrients. One major function of the hindgut is to reabsorb the fluids secreted into the upper digestive tract and (in animals that have a cloaca) excreted in the urine. It also serves as the principal site for the microbial production of nutrients in the herbivorous reptiles and birds and in most herbivorous mammals. Thus, the hindgut tends to be longest in animals that need to conserve water in an arid environment, and it has a larger capacity in most herbivores.
Musculature
The digestion of food, the absorption of nutrients, and the excretion of waste products require the mixing of ingesta with digestive enzymes and the transit of ingesta and digesta through the digestive tract. In all vertebrates (except the cyclostomes), the contents are mixed and moved by an inner layer of circular muscle and an outer layer of muscle that runs longitudinally along the tract (Fig. 7). See also: Excretion; Muscle; Muscular system
The initial act of swallowing and the final act by which waste products are defecated from the digestive tract are effected by striated muscle. This type of muscle is characterized by rapid contraction and is controlled by extrinsic nerves. The esophagus of fish and varying lengths of the mammalian esophagus are enveloped with striated muscle. However, the esophagus of amphibians, reptiles, and birds and the entire gastrointestinal tract of all vertebrates are enveloped by smooth muscle. This smooth muscle contracts more slowly, and its rate of contraction is partly independent of external stimulation.
Nerve and endocrine tissues
Deglutition and defecation are under the voluntary control of the central nervous system. However, the remainder of the digestive system is subject to the involuntary control of nerves that release a variety of neurotransmitting or neuromodulating agents that either stimulate or inhibit muscular contractions and the secretions of glands and cells. The motor and secretory activities of the digestive system are also under the control of a wide range of other substances produced by endocrine cells that are released either distant from (hormones) or adjacent to (paracrine agents) their site of action. Although there are some major variations in the complement and activities of the neurotransmitters, neuromodulators, hormones, and paracrine agents, their basic patterns of control are similar. See also: Endocrinology; Hormone
Physiology
The major physiological activities of the digestive system are motility, secretion, digestion, and absorption. Each activity can be affected by diet and, in the cold-blooded species, is reduced with a decrease in body temperature.
Motility
The mastication of food and the movement of ingesta and digesta through the digestive tract are controlled by the motor activity of muscular contractions. The pressure of food against the palate and back of the mouth stimulates a nerve reflex that passes through a deglutition center in the brain. This reflex closes the entrance into the respiratory system and stops respiration, thereby preventing the inspiration of food into the lungs, and initiates muscular contractions that pass food into the esophagus. The food (bolus) is then passed down the esophagus and into the stomach by a moving wave of muscular contractions (peristalsis) accompanied by inhibition of the esophageal sphincters.
The multicompartmental forestomach of ruminants undergoes a continuous series of complex, repetitive contractions that are controlled by the central nervous system. However, the gastric motility of most species and the intestinal motility of all vertebrates are controlled partially by the intrinsic characteristics of their smooth muscle cells. The result is production of either stationary (mixing) contractions of the stomach and intestine or a series of peristaltic contractions that carry digesta on through the tract.
Digestion
The diet of vertebrates contains carbohydrates, lipids, and proteins, which provide the energy and building blocks for growth, maintenance, and reproduction, and the vitamins that are required in small amounts for the regulation of metabolic processes. The lipids, proteins, and most of the carbohydrates require digestion for the release of nutrients that can be absorbed. Digestion is accomplished by enzymes produced by the digestive system (endogenous enzymes) or by bacteria that are normal residents of the digestive tract. Plant and animal starches are converted to oligosaccharides (short-chain structures) and disaccharides by amylase, which is secreted by the salivary glands of some species and the pancreas of all vertebrates. The end products of starch digestion, plus the dietary disaccharides, are converted to monosaccharides by enzymes in the brush border of the absorptive epithelial cells lining the small intestine. Vertebrates do not produce enzymes capable of digesting the structural polysaccharides of plants. See also: Carbohydrate; Carbohydrate metabolism; Oligosaccharide; Polysaccharide; Vitamin
Lipids are digested into alcohols, monoglycerides, and fatty acids by lipases and esterases, which are secreted predominantly by the pancreas. However, the lipases are water-soluble enzymes that can attack their substrate only at a lipid–water interface. Therefore, the lipids must be emulsified in order to provide the surface area required for efficient digestion. Emulsification is accomplished by the release of bile salts secreted by the liver and released into the midgut. See also: Lipid metabolism
Dietary protein is first broken down into long chains of amino acids (polypeptides) by gastric pepsin and pancreatic trypsin. The polypeptides are then attacked by other pancreatic proteases (chymotrypsin, carboxypeptidase, and elastase) to form tripeptides, dipeptides, and amino acids. All of these enzymes are secreted in an inactive form to prevent the self-digestion of the secretory cells prior to their release. Pepsin is activated by the acidity resulting from the secretion of hydrochloric acid (HCl) into the stomach, and trypsin is activated by an enzyme (enterokinase) that is secreted by the intestinal epithelium. Tripeptides and dipeptides are digested into amino acids by enzymes in the brush border and contents of midgut absorptive cells. Nucleic acids are digested by pancreatic ribonucleases into pentose sugars, purines, and pyrimidines. See also: Amino acid; Nucleic acid; Protein
Importance of indigenous bacteria
Substantial numbers of bacteria can be found in all segments of the gastrointestinal tract, but the highest numbers are present in those segments in which digesta are retained for prolonged periods of time at a relatively neutral pH. These indigenous bacteria (often termed the microbial flora, microbiota, or microbiome) help protect the animal from pathogenic microorganisms by stimulating immunity and competing for substrates. They also convert dietary and endogenous substances that are not digested by endogenous enzymes into absorbable nutrients. Many species of indigenous bacteria can ferment sugars, starches, and structural carbohydrates into short-chain fatty acids. The short-chain fatty acids, which are predominantly acetic, propionic, and butyric acids, are readily absorbed and serve as an additional source of energy. These bacteria also synthesize microbial protein and the B-complex vitamins that may be useful to their host. See also: Bacteria; Bacterial physiology and metabolism; Human microbiota; Microbiome
Absorption
The epithelial cells that line the gastrointestinal tract are closely attached to one another at their lumen-facing border by tight junctions, which are relatively impermeable to most substances other than water. Therefore, the major restriction for the absorption of most substances from the lumen into the blood is the apical and basolateral membranes of these cells. Lipid-soluble substances can be transported across the apical cell membranes by passive diffusion down their concentration gradient. The short- and medium-chain fatty acids that result from lipid digestion in the small intestine pass directly into the blood. However, the monoglycerides and long-chain fatty acids are resynthesized into triglycerides by the epithelial cells in the midgut and incorporated into small spheres (chylomicrons), which are transported across the basolateral membrane into the lymphatic system. Fat-soluble vitamins, long-chain alcohols, and other lipids also appear to be incorporated into chylomicrons, and they too enter the lymphatic system. See also: Cell membranes; Triglyceride (triacylglycerol)
The intestinal cell membranes are relatively impermeable to the passive diffusion of water-soluble monosaccharides, amino acids, vitamins, and minerals that constitute a major portion of the required nutrients. These nutrients are selectively transferred across the intestinal cell membranes by carrier-mediated transport. Membrane carriers combine with the nutrient at one membrane surface and pass it across the membrane for release at the opposing surface. Some simply facilitate the diffusion of a substance down its concentration gradient; others are capable of transporting a nutrient against its concentration gradient, which requires either a direct or indirect investment of cellular energy.
Above all, the nutrient that is required in the largest quantity for the digestion, absorption, metabolism, and excretion of waste products is water. Because it readily diffuses across cell membranes down its concentration gradient, the net secretion or absorption of water is determined by the net secretion or absorption of all other substances. Sodium, chloride, and bicarbonate are the principal ions that are present in the extracellular fluids that bathe the body cells of all vertebrates and that are transported across cell membranes. Therefore, the transport of these electrolytes is the major driving force for the secretion or absorption of water. See also: Electrolyte; Water

